Ascomycota
Back to main indexJump to section:
Introduction
The Ascomycota, ascomycetes in the vernacular language, form the group of Dikarya which contains the greatest number of species, since approximately 65% of the species of all described fungi belong to this lineage. The shared character of the group, recognized as early as the 19th century, is the fact that their meiospores are produced inside a sac called ascus, hence their name ascospores (Figure 140). Phylogenetic analyzes confirm that the group is monophyletic and show that these fungi have diversified into three independent lineages: Taphrinomycotina, Saccharomycotina, and Pezizomycotina (Figure 141). That of Taphrinomycotina diverged first from the other two. The fungi of Taphrinomycotina and Saccharomycotina can be considered simple in form and function. On the other hand, the class of Pezizomycotina contains species with complex characters.
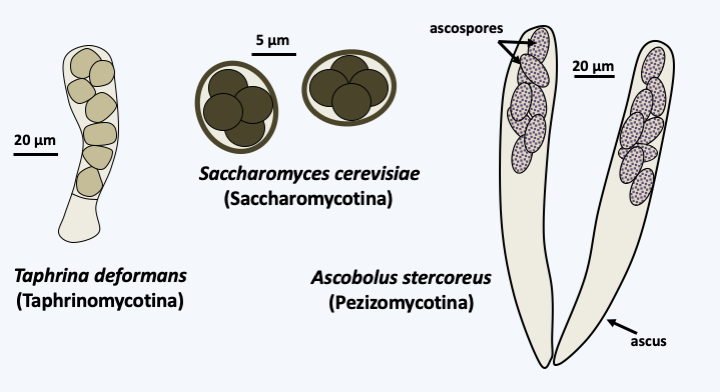
Figure 140.
Asci and ascospores.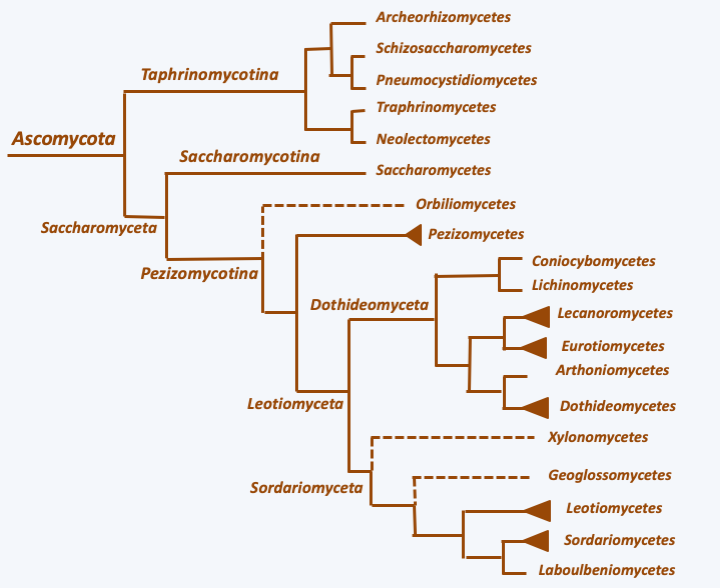
Figure 141.
Phylogeny of Ascomycota.Taphrinomycotina
The Taphrinomycotina, formerly called Archiascomycetes, form a group of “primitive” Ascomycota comprising a few dozen species, but with very varied biology. Currently, five classes are distinguished.
Taphrinomycetes
Taphrinomycetes are parasites of plants. In nature, they are mainly found in the leaves as dikaryotic mycelia, a condition also exhibited by Basidiomycota, but not by other Ascomycota where the dikaryotic state is confined to a fruiting body. This mycelium cannot be cultivated in an axenic medium. Monokaryotic strains are yeast-like and can be cultured. The type species of the class is Taphrina deformans, a pest of peach and almond (Figure 142). Its genome has been sequenced. It encompasses 13.5 Mb and contains around 6,000 genes, which makes it more like yeast genomes than those of mycelial species which have genomes twice as large or even larger. The life cycle (Figure 143) is essentially sexual.

Figure 142.
Symptoms created by Taphrina deformans on a peach tree.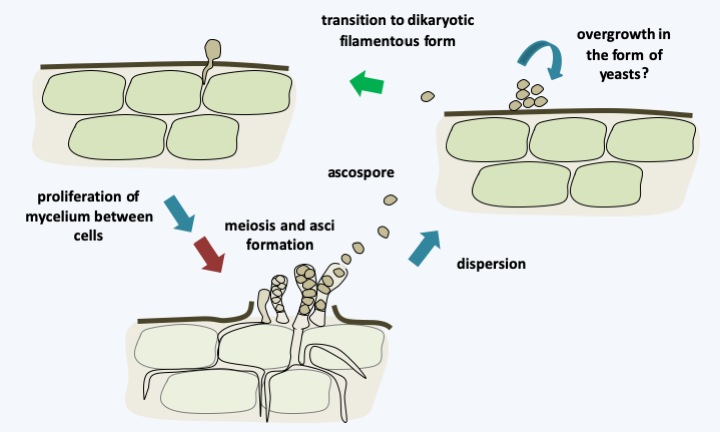
Figure 143.
Cycle of Taphrina deformans. The asci are formed directly on the mycelium and exposed to the external environment. They discharge their spores violently like some Pezizomycotina. The release of ascospores and their germination results in the laboratory haploid saprotrophic yeast colonies. The inoculation of these yeasts allows the plants to be infected. Taphrina deformans is homothallic and infection with yeasts from a single cell is sufficient to complete the cycle. This ends after proliferation of the dikaryotic mycelium between host cells by meiosis and post-meiotic mitosis leading to the formation of 8-ascospore asci.Neolectomycetes
There are only three species of Neolectomycetes all belonging to the genus Neolecta. These fungi form multicellular fruiting bodies (Figure 144). These appeared by convergent evolution. Indeed, although the ontogeny of these fruiting bodies is unknown, the manufacture of the asci that they bear occurs in a different way from that of Pezizomycotina. In particular, there is no hook and the two nuclei that will merge are attached by a structure visible under electron microscopy. Analysis of the pore between the cells of the mycelium, the only one performed to date for a Taphrinomycotina, shows a simple structure different from that found in Pezizomycotina. Unfortunately, these fungi cannot be cultivated in a Petri dish, due to the fact that Neolecta are probably mycorrhizal fungi.
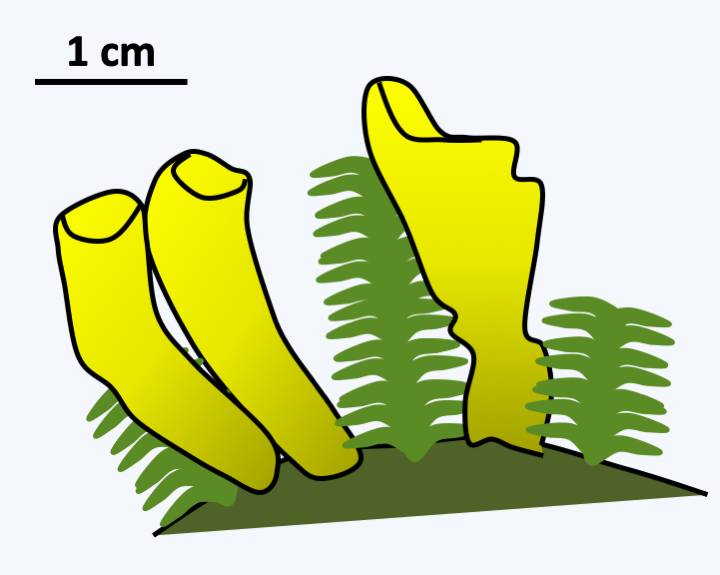
Figure 144.
Carpophores of Neolecta irregularis.Archaeorhizomycetes
Species in the Archaeorhizomycetes class have only recently been discovered and have long been known only from environmental DNA sequences. There are hundreds of species that seem to live associated with the roots of plants. Not forming mycorrhizae, these fungi are thought to be saprotrophs feeding on root exudates. The high frequency with which they are detected indicates that these fungi are ubiquitous in soils, especially in tundra, and probably form a considerable biomass. It is only very recently that their cultivation has allowed them to be better studied. The slowness of their growth (a few mm per month) does not allow us to conclude on their trophic mode. They are so far not known to have a sexual cycle, or even asexual. Bulges of hyphae are however present and could serve as chlamydospores.
Schizosaccharomycetes
The class of Schizosaccharomycetes contains only four species of saprotrophic yeast dividing by fission. The best known member is the yeast Schizosaccharomyces pombe, commonly known as the fission yeast, which is used in Africa to make beer. Nowadays it is used in many research laboratories for studies of various aspects of cell biology. In fact, we can easily select mutants in this yeast and carry out genetic analysis. It has served in particular as a model for the study of the cell cycle. It reproduces by binary fission. The species has a haplobiontic life cycle with two mating types (Figure 145). Its genome sequence is available. It is distributed over three chromosomes, and is the genome of a non-parasitic eukaryotic cell that contains the smallest number of genes; the current estimate is 4,824.
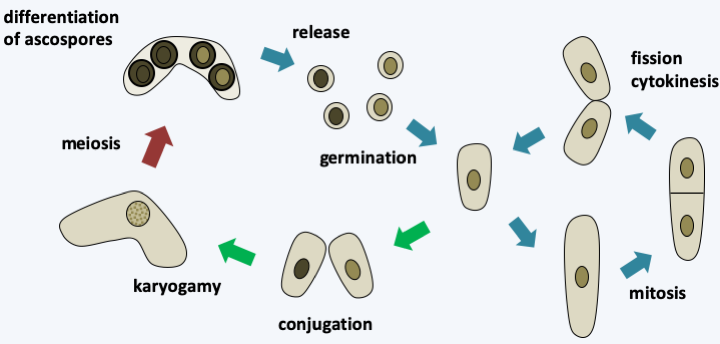
Figure 145.
Cycle of Schizosaccharomyces pombe. The cycle is haplobiontic in nature, but it is possible in the laboratory to grow diploids which then divide like haploids. Yeast conjugates when nitrogen deficiency occurs and under normal conditions diploids immediately perform meiosis to give 4-spore asci.Pneumocystidiomycetes
The last class of the Taphrinomycotina, the Pneumocystidiomycetes, contains species that have long been classified as parasitic protozoa. Indeed, the Pneumocystis are commensals or obligate parasites of the lungs of mammals. Usually the infection is asymptomatic, but becomes serious if the immune system is weak. Molecular data shows that there are several species that each specifically infect a host. There could even be as many species of Pneumocystis as there are species of mammals! Pneumocystis jirovecii specifically infects humans (Figure 146). It is probably the most common opportunist in AIDS patients. This fungus is present all around the world and enters the body via the respiratory tract. However, the infectious form that spreads the disease is still unknown. Once in the lungs, it mainly grows inside the alveoli and causes pneumonia if the immune system is weak. Unfortunately, we do not know how to cultivate these fungi in vitro, which makes their study difficult. The genome, made up of about fifteen chromosomes, is small (8 Mb) and has a high level of T and A nucleotides (65%). Unusually for Ascomycota but common for Basidiomycota, the genes are rich in introns.
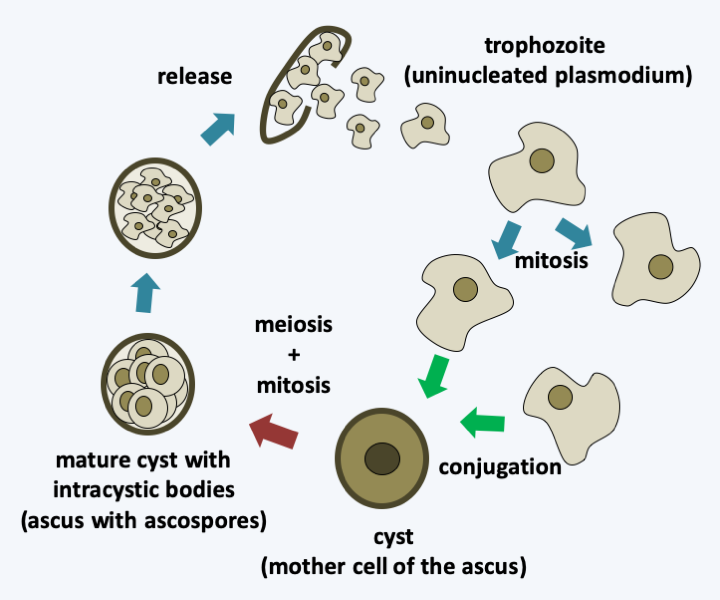
Figure 146.
Cycle of Pneumocystis jirovecii. Haploid trophozoites divide by mitosis. The conjugation of two trophozoites leads to the differentiation of a cyst in which meiosis occurs and postmeiotic mitosis produces eight intracystic bodies. These will be released by rupture of the wall of the cyst. The terminology used is that for parasitic protozoa; in parenthesis that for mushrooms which should be used.Saccharomycotina
The Saccharomycotina, formerly the Hemiascomycetes, represent only 1% of the species of fungi. Despite this, this group contains many species important to human industry, including the bread / wine / beer yeast, Saccharomyces cerevisiae. As a result, Saccharomycotina serve as study models in many laboratories and as a factory for the production of proteins and metabolites in industry. In particular, Saccharomyces cerevisiae has been used extensively to analyze the cellular physiology of the micropores or plasmodesmata of the eukaryotic cell (Box 16). Currently, Saccharomycotina are used to understand the evolution of genomes, resulting in the availability of the greatest number of complete genome sequences for a defined group of eukaryotes (see Figure 112. There are at least 600 species, but their diversity must be greatly underestimated, because a recent inventory of the diversity of yeasts present in the intestines of tropical beetles has made it possible to double the number of Saccharomycotina species!
Box 16. The Saccharomycotina yeasts Saccharomyces cerevisiae and Candida albicans
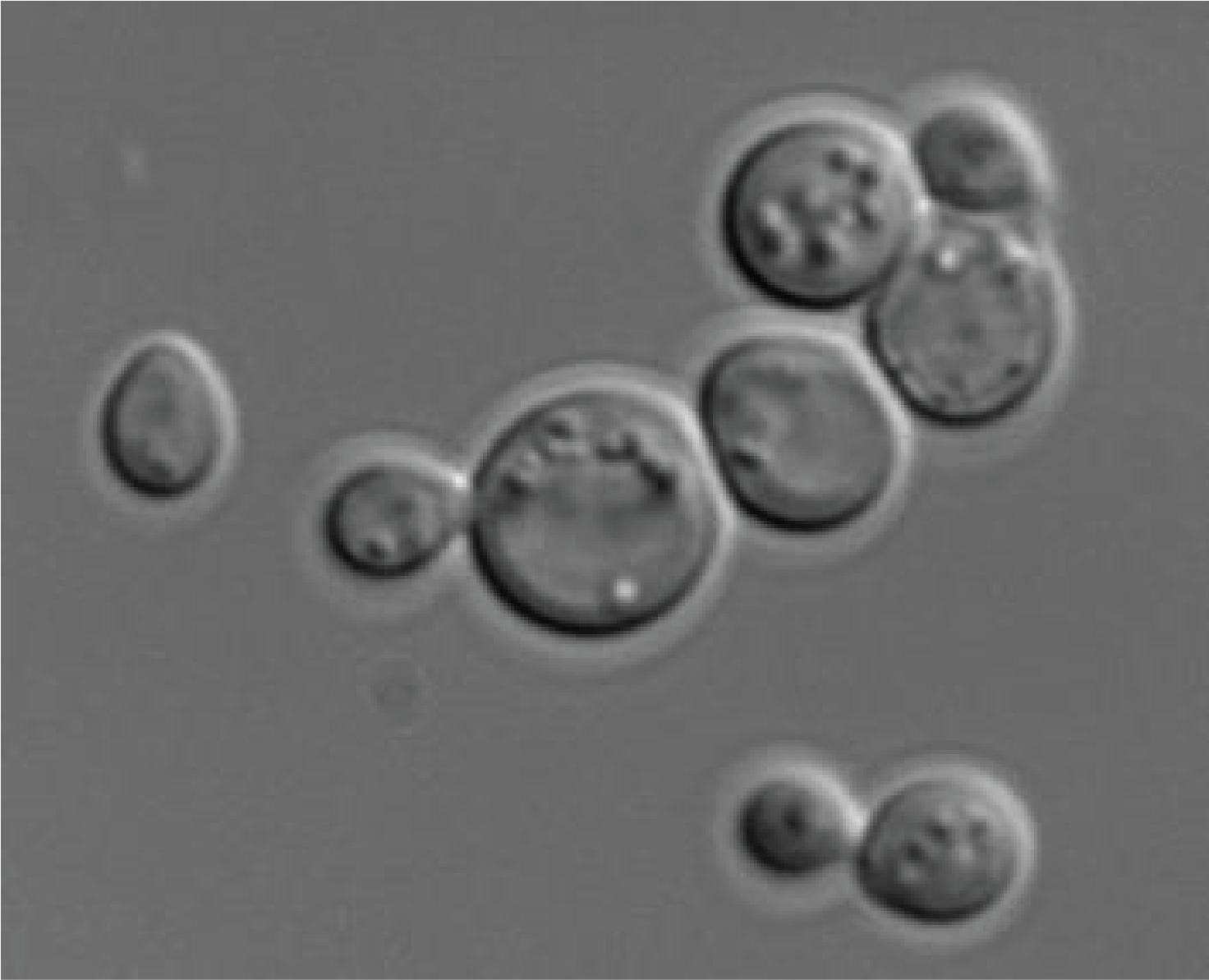
Two species of Saccharomycotina are especially important to human societies, which has led to intensive study of their biology: Saccharomyces cerevisiae and Candida albicans.
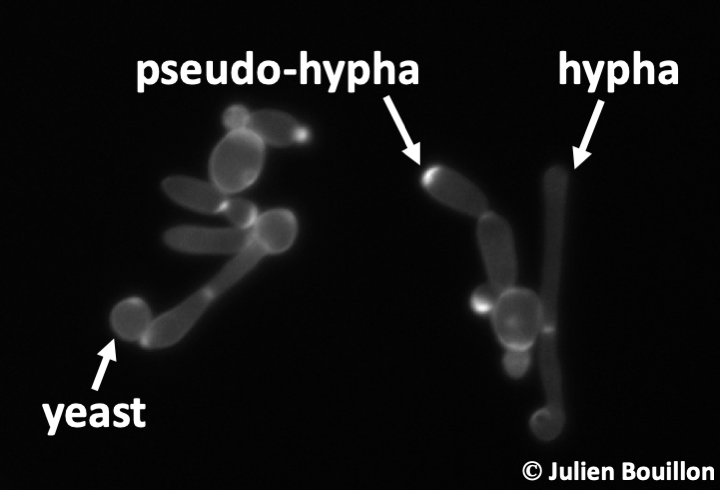
Saccharomyces cerevisiae has been cultivated by humans for over 9,000 years to make wine in Asia Minor, over 6,000 years to ferment beer in Babylon, and over 2,000 years to make bread in Egypt. It is found naturally in the sap secretions emitted by trees and on decaying fruit where it feeds on the present sugars. Nevertheless, the easiest way to harvest it is to visit the fermentation tanks of vineyards! Saccharomyces cerevisiae is found as a spheroidal uninuclear cell measuring about 20 microns. Its size is dependent on several factors: the strain, its ploidy, and the age of cell, because it divides by budding. Under certain conditions of stress, it exhibits growth with pseudo-hyphae and can sometimes be invasive. Generation time under good conditions is around 80 minutes. Its life cycle is haplodiplobiontic with two mating types referred to as mat a and mat α. Growth in the absence of oxygen results in the exclusive use of fermentation and most enzymes in respiration are then repressed. Fermentation is sufficient to ensure the survival of the cell, so Saccharomyces cerevisiae is a facultative aerobic organism. When the strains are grown under aerobic conditions in the presence of a high concentration of glucose, it begins by fermenting the sugars into ethanol and CO2 with consumption of oxygen. If oxygen runs out, a change in metabolism happens, switching instead to anaerobic fermentation of glucose with the production of ethanol until its complete consumption, followed by entry into the stationary phase. It is this latter metabolism that is responsible for using yeast to make bread, wine, and beer.
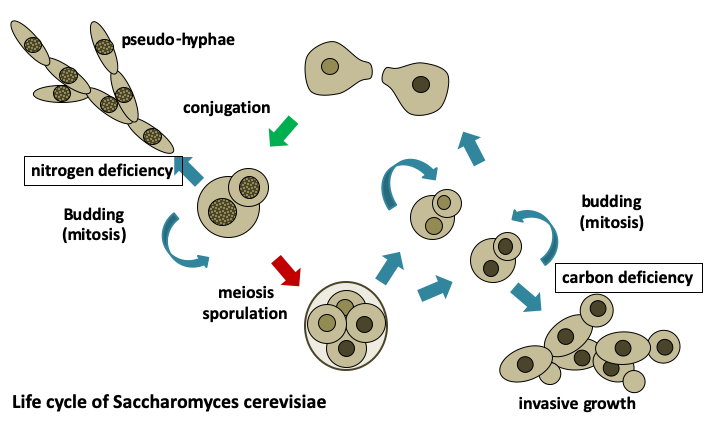
During the last two centuries, Saccharomyces cerevisiae has supported researchers as a model organism. Pasteur in the 19th century used it to show that fermentation processes are due to microorganisms and that each type of fermentation is associated with particular microorganisms, thus laying the foundations for modern microbiology. In 1897, Buchner showed that cell-free extracts of yeast could convert glucose into ethanol and CO2, showing that energy metabolism can be carried out by inert matter, and that the responsible products could be purified, thus opening the way for the field of biochemistry. Saccharomyces cerevisiae was introduced for genetic studies during the 1930s and during the 1970s and 1980s it was established as the major model for molecular and cellular biology studies in eukaryotes. This culminated with the complete sequencing of its genome in 1996 and the systematic deletion of all of its genes in 2000. It was the first complete sequence of a eukaryotic organism. It is undoubtedly the eukaryotic organism best known to scientists. When searching the NCBI 'PubMed' database with the term Saccharomyces, over 115,000 citations are returned. The number of scientific articles published on this organism is approximately 4,000 per year!
Candida albicans is currently the fungus responsible for the greatest number of yeast infections in humans. Its detection is difficult, since this species is a normal part of the microflora of the intestine. Unbalanced proliferation of the fungus gives rise to disease. Systemic infections affect immunocompromised patients and are often fatal. They begin with the invasion of the wall of the digestive tract where the yeast attaches. When present in large quantities, this can lead to malnutrition. The yeast can then invade other tissues. It is postulated that all humans have had problems with these yeasts at least once in their life. Candida now constitutes about 10% of nosocomial (hostpital-derived) infections and pose serious problems for immunocompromised patients. Besides yeast forms, they produce pseudo-hyphae or true hyphae. They can also form chlamydospores. The transition between yeast and hypha is important in the infectious process; the yeasts being adapted rather for dissemination and the hyphae for penetration into the host. This yeast can also escape macrophages and therefore persist in the body.
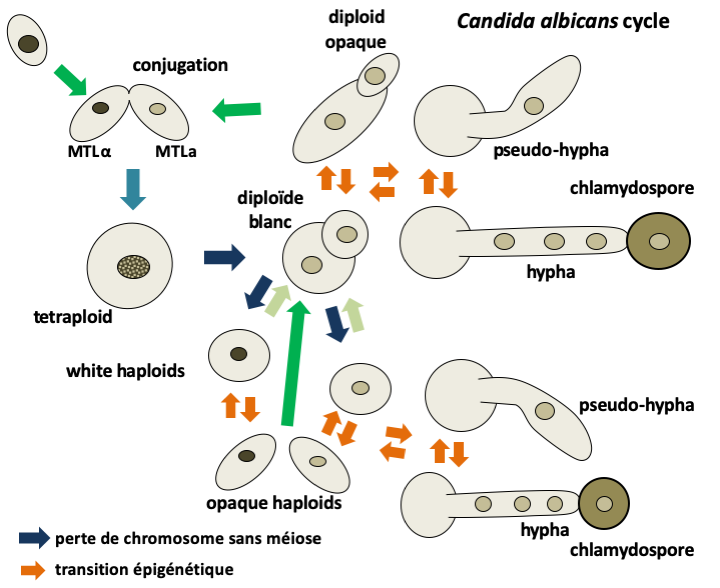
Candida albicans reproduces only asexually and never goes through meiosis. However, a process related to sexuality has been described in this fungi. The strains are generally diploid but with many variations of karyotypes including aneuploidies. A few rare haploid strains have been identified; they derive from diploid strains by loss of chromosomes and can directly give back diploids, probably after a defect in separation of daughter nuclei after mitosis. They have the same vegetative differentiation capacities as diploid yeasts. The two mating type factors MTLa and MTLα, homologous mat a and mat α of Saccharomyces cerevisiae, are present in the Candida albicans genome. Most diploid strains have both mating type factors and are unable to conjugate. They have a constant morphology which leads to a 'white' appearance of the colonies. On the other hand, when one of the mating type factors is absent, Candida albicans becomes capable of conjugating with another strain, if it is homozygous for the complementary mating type. Conjugation can occur between haploids to give back a diploid or between homozygous diploids of complementary mating types to give rise to a tetraploid! However, not all homozygous cells engage in conjugation with the same frequency. In fact, the colonies of homozygous strains can adopt two morphological forms: a white one resembling heterozygous diploid strains and a so-called opaque form having more elongated cells. The frequency of conjugation is a hundred times higher in the opaque form. On the other hand, white cells are much more resistant because their wall is thicker. The passage between the white and opaque forms is controlled by an epigenetic mechanism involving the self-activation of a transcription factor. It is probable that this is an adaptation to the hostile environments that Candida albicans encounters in the body: the white form allows it to resist and the opaque form to conjugate with neighboring cells and therefore to be able to recombine genetic combinations to better adapt to its environment. After conjugation, the return of the tetraploid to the diploid state is poorly understood, but occurs through random loss of chromosomes as during the transition from the diploid state to the haploid state. This random reassortment of chromosomes is referred to as the parasexual cycle.
Genomic analyzes show that these organisms derive from more complex fungi by reductive evolution. They do not form multicellular fruiting structures and most are yeasts which divide by budding. However, some species produce pseudohyphae under certain conditions, even true hyphae such as Geotrichum candidum, Ashbya gossypii or Yarrowia lipolytica. These hyphae are nevertheless simpler than those of other fungi with in particular the absence of a central pore and the presence of micropores (Figure 147). Most species are saprotrophs that consume simple sugars or lipids, but they can also be found as commensals of vertebrates or insects. Ashbya gossypii is a pest of cotton. In the Saccharomycotina, we find Candida, of which about ten species are opportunistic pathogens in humans. Candida causes skin or systemic infections that can be transmitted by different routes including the sexual route. The most important species today is Candida albicans, which is the most common pathogenic fungus in humans (Box 16).
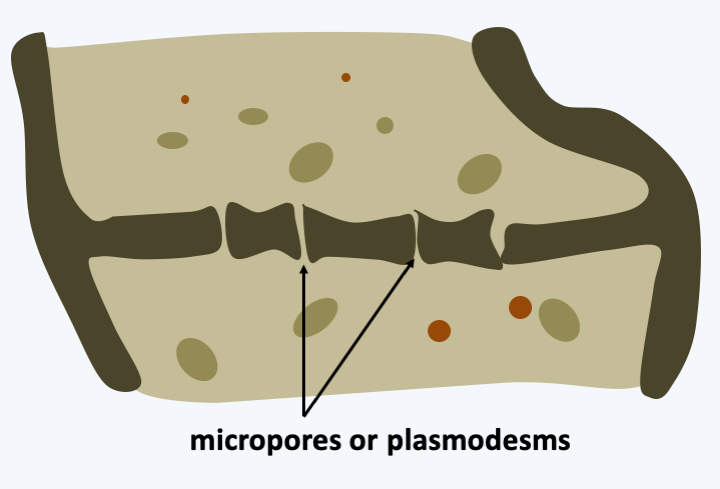
Figure 147.
Micropores of Saccharomycotina.Pezizomycotina
The Pezizomycotina, formerly Euascomycetes, represent the vast majority of Ascomycota with several tens of thousands of species described, or 60% of all known fungi. They have invaded most environments: water, soil, plants and animal intestines… They have evolved in recurrent ways towards all styles of life. Half of the species described live in association with algae to form lichens. However, this proportion does not reflect the true proportion because most of the lichen species are already described while those of the other trophic groups are only partially known. Saprotrophs are very numerous and well known to spoil our food. A few are ectomycorrhizals, many more are parasites or symbionts of plants or animals. Most species exhibit characteristics typical of “complex” fungi. They have an integrated filamentous thallus with efficient anastomoses and so-called Woronin-seeded septal pores that connect the cells together (Figure 148). Some species may switch to a mode of growing as a yeast. Many Pezizomycotina build multicellular fruiting bodies often of sizes less than 1 mm but sometimes of large sizes, like the elfin saddles (Figure 149). These fructifications served as a basis for classification until the end of the 20th century (Box 17). Molecular phylogenies have since shown that fruiting bodies have evolved convergently multiple times. Besides ascospores, Pezizomycotina are capable of producing numerous mitospores which can ensure asexual dissemination. There are several mechanisms that make it possible to produce these mitospores, such as the fragmentation of the mycelium or the differentiation of certain cells. The most common are the conidia or conidiospores which differentiate at the tip of a particular hyphae, the conidiophore (Box 17). Conidiophores can be isolated or grouped into synemata (sing. synema). They are either directly exposed to the outside or protected in structures called acervuli and pycnidia (Box 17). Most species therefore have a sexual cycle and an asexual cycle.Figure 150shows the generic life cycle, but there are many variations across species. Molecular phylogenies divide the Pezizomycotina into thirteen classes, six of which are important both for the number of species they possess and for their ecological roles.
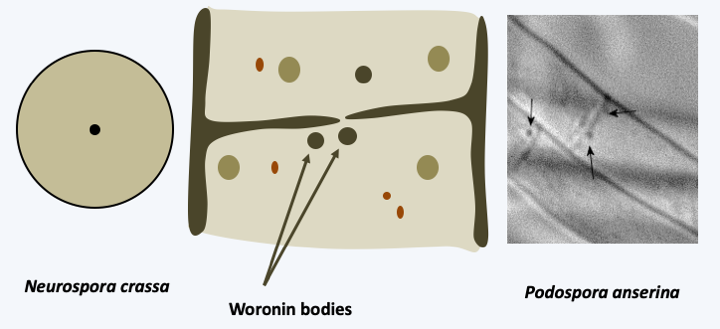
Figure 148.
Woronin bodies of Pezizomycotina. Woronin bodies are modified peroxisomes containing only one protein. They are positioned near the central pore, seen from the front left. They pour out their contents to occlude the pore if one of the two items suffers an injury.
Figure 149.
Some fruiting bodies of Pezizomycotina. The fruiting bodies of Helvella crispa or Ascocoryne sarcoides are clearly visible to the naked eye, while those of Podospora anserina and Erysiphe flexuosa are not.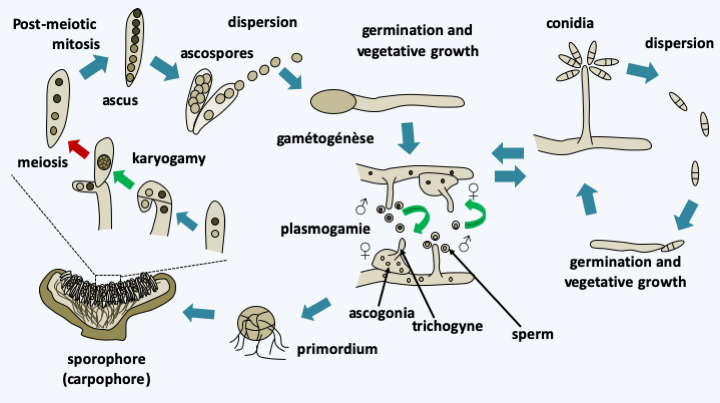
Figure 150.
Generic life cycle of Pezizomycotina. Most species are able to complete asexual and sexual cycles. The asexual cycle most often involves the production of conidia that disperse easily in the air. The sexual cycle begins with the differentiation of male and female gametes. The male gamete is either a cell resembling a conidium, the spermata, or a particular hypha, the antheridium. The female gamete is a large cell with a particular morphology, the ascogonium. The fungus can be homothallic or heterothallic of the bipolar type. Plasmogamy between the ascogonium trichogyne and the male gamete leads to the dikaryon. In all Pezizomycotina, the characteristic dikaryotic stage occurs in fruiting and is maintained thanks to the cellular mechanism described in figure 131. It allows the generation of numerous meiosis from a single fertilization. Karyogamy takes place in the parent cell of the ascus and is immediately followed by meiosis and ascospore formation. Note that in some species it is not the apical cell which sees the karyogamy of the two nuclei but the basal cell which reforms. The presence of post-meiotic mitosis results in eight-spore asci. In some species, several post-meiotic mitoses may occur resulting in asci with 16, 32, 64 or 128 ascospores. Likewise, in some cases, two nuclei resulting from meiosis or mitosis can be found in the same ascospore, resulting in the formation of asci with four dikaryotic ascospores. Usually, once the ascospores are ripe, they are ejected from the fruiting body at a very high speed. It is in fact the fastest flight currently known in nature with accelerations of 180,000 g giving speeds of up to 25 m/s. Note that to reach such speeds, it is not necessary to imagine complex mechanisms because a conventional turgor pressure is sufficient to provide propulsion. This type of ascospore expulsion mechanism partly explains in these fungi the position in the fruiting body of the hymenium which is most often directed upwards.Box 17. Fruiting bodies of the Pezizomycotina
As in Basidiomycota, the shape of the sexual fruiting bodies has long served as the basis for the classification of Pezizomycotina. Classically, mycologists distinguished between discomycetes producing apothecia, pyrenomycetes developing perithecia or pseudothecia and plectomycetes producing cleistothecia or gymnothecia.


Apothecia are fruit bodies where the hymenium is open to the outside. As with Ciboria batschiana, they are most often cup-shaped over a small foot with the hymenium positioned on the upper surface. However, in some species they take on more complex forms (Figure 149, Helevella crispa). Their size varies from a few tens of micrometers to more than ten centimeters in diameter. The presence of this type of fruiting in the majority of Pezizomycotina, including Orbiliomycetes and Pezizomycetes which are the lineages that diverged the earliest from the rest, indicates that apothecia are the ancestral type from which the others are derived. For example, the truffles that we consume, the black truffle from Périgord and the white truffle from Piedmont, derive from apothecia by closure and internal folding of the hymenium.
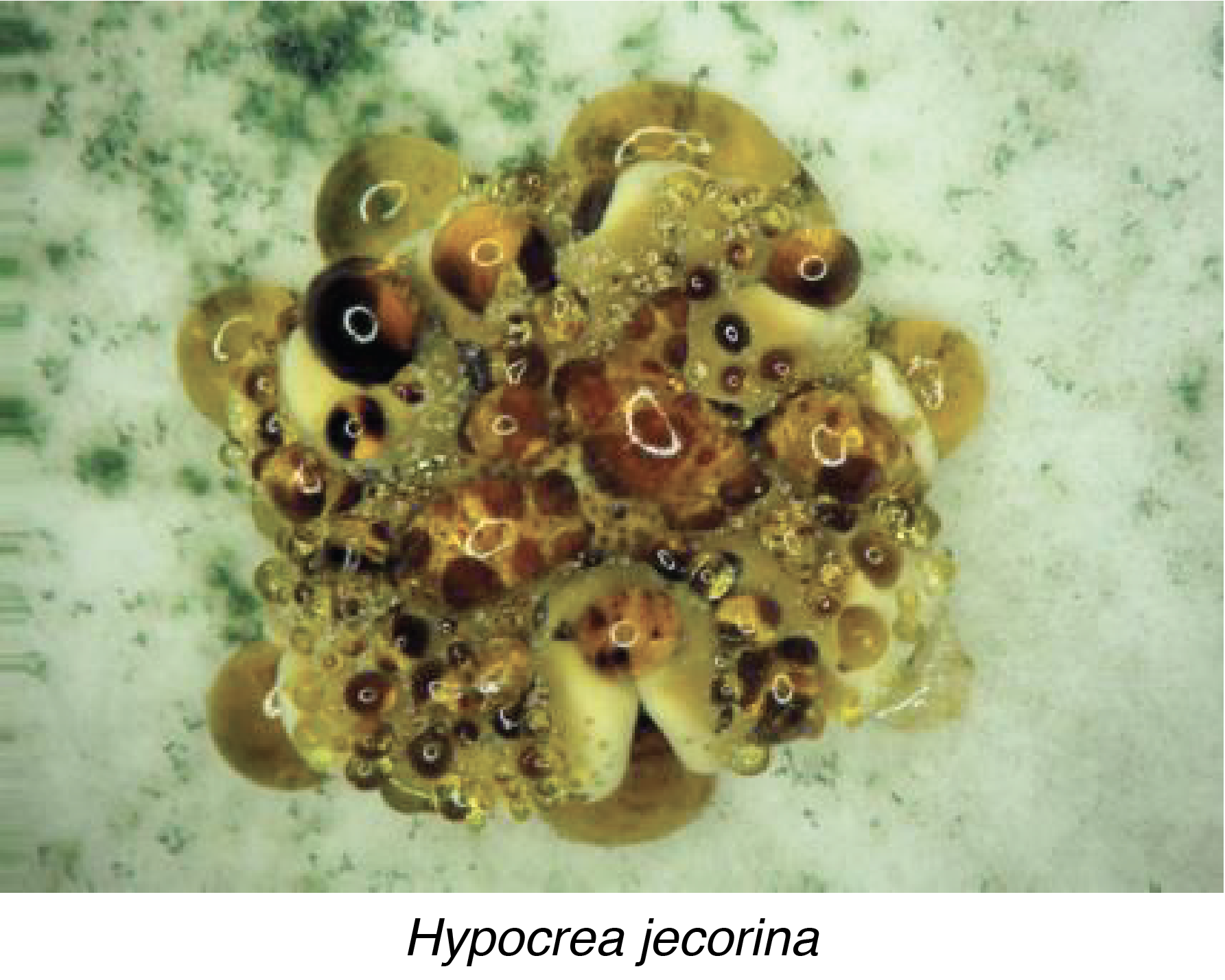
The perithecia have a protected hymenium inside a peridium that is pear-shaped and about 1 mm high. The perithecium may be glabrous, or covered with 'hairs' as in Lasiosphaeria ovina. The hymenium lines the bottom of the structure, which allows the asci to be oriented vertically. The ascospores can then escape through the ostiole positioned at the top of the perithecium. Pseudothecia have a structure similar to that of perithecia, but differ in their ontogeny. This is because perithecia develop around a female gamete after it is fertilized. This type of development is called ascohymenial. On the contrary, the pseudothecium differentiates first before the female gamete which is therefore fertilized once the pseudothecium has completed its development, which is said to be ascolocular. Pseudothecia are also often associated with 'bitunicate' asci. These asci are surrounded by two walls that crack sequentially to allow the expulsion of ascospores. As in Hypocrea jecorina, pseudothecia and perithecia can be grouped together in clusters of hyphae called stromata (sing. stroma).
Cleistothecia are completely closed fruiting bodies. Note that truffles are sometimes considered cleistothecia, however, the majority of mycologists reserve this name for small fruiting bodies without a hymenium arranged in a layer. Indeed, in a typical cleistothecia, asci are no longer arranged as a differentiated hymenium, while truffles still have a well-defined hymenium. Ascospores are no longer actively expelled outside the cleistothecium and therefore serve more for persistence than dispersal, as in Aspergillus nidulans. Gymnothecia are similar in structure to cleistothecia but have a looser peridium made up of interlocking hyphae.

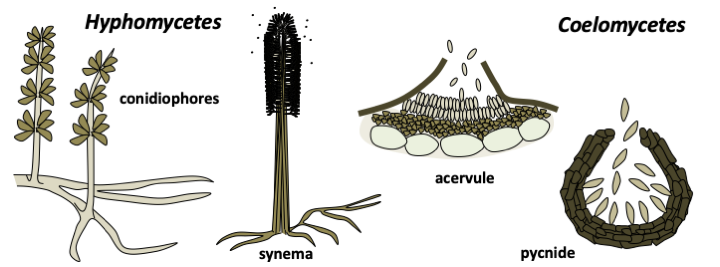
Many Pezizomycotina are only known by their asexual (anamorphic) sporophores. Formerly, they were classified in the class of Deuteromycetes with all the other fungi known only by their form of asexual dispersion. This artificial class no longer exists and the sexual (teleomorphic) forms of many species are being identified with increasing frequency. However, it is often easy to recognize a fungus by analyzing the anamorph. Traditionally, two main types are defined: Hyphomycetes forming mitospores directly on the mycelium and Coelomycetes producing spores in cavities.
Orbiliomycetes
The class of Pezizomycotina that seems to be the earliest diverging is the Orbiliomycetes. This placement is supported by some molecular phylogenies, but not all. The ultrastructure of the asci and septal pores appears to be simpler than in other Pezizomycotina, supporting the hypothesis of early divergence from the rest of the clade. There are a few hundred species. They are saprotrophs living mainly in soils. Their sexual cycle is not well known because teleomorphs are rarely observed and produce very small apothecia (fruiting bodies). Most species trap nematodes to supplement their diet. This is the case with Arthrobotrys oligospora which captures nematodes and other small animals via special snare-shaped hyphae (Figure 151). The prey is then digested by extracellular enzymes.
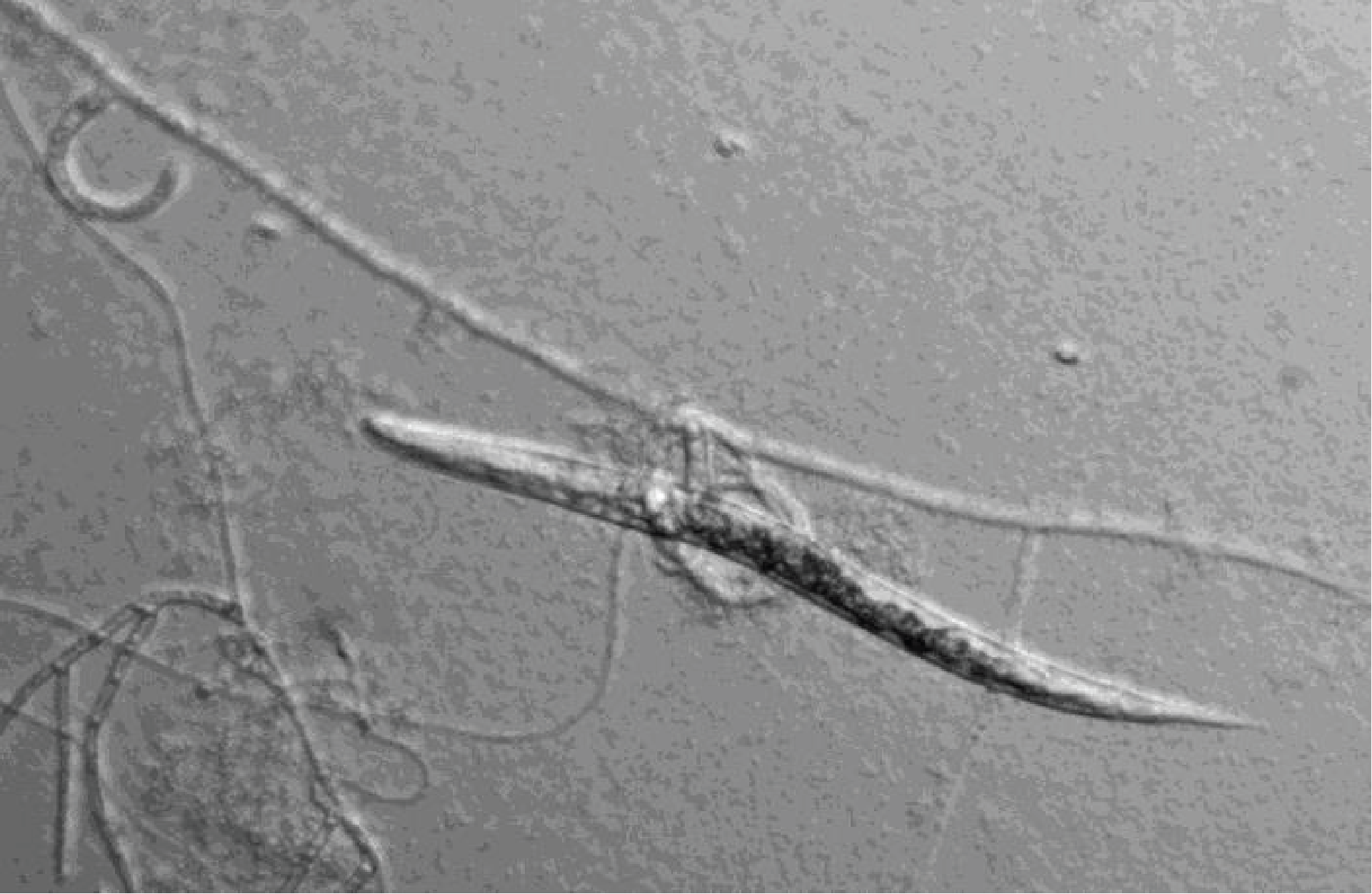
Figure 151.
Nematode trapped by Athrobotrys oligospora.Pezizomycetes
The other candidate for being the most divergent class is that of the Pezizomycetes. This lineage includes fungi which have apothecia-type fruiting bodies, very often of large size. The most famous members have a culinary interest, such as truffles, morels and the orange-peel fungus (Figure 152). More than 1,200 species of Pezizomycetes are currently described. In this group, mainly saprophytes are found, but also some species forming ectomycorrhizae such as the truffle with oak and some other trees. Other species live associated with plants without us always clearly knowing the relationships they maintain. For example, morels are saprotrophic but seem to be able in certain cases to engage in mycorrhizae. Others are clearly pests of plants, like Neottiella. Despite the gastronomic interest, the fruiting of these mushrooms is so far poorly controlled. For example, the truffle requires the presence of trees to fruit and the required signal is unknown. It is also in this group that the Ascobolus are found, which are used as models in some laboratories or in biology classes. Ascobolus have microscopic fruiting bodies and are common on herbivore droppings.

Figure 152.
Two Pezizomycetes. On the left, the orange peziz, Aleuria aurantia, and on the right the conical morel Morchella conica. Both species are edible.Sordariomyceta
The other Pezizomycotina are divided into two major lineages: the “Sordariomyceta” and the “Dothideomyceta” (these terms are not formal taxonomic groups). The Sordariomyceta group together five classes, two of which are interesting for their low number of species but important roles in ecosystems. The class of Geoglossomycetes contains only a few species. These fungi are saprotrophic and form large apothecia in the shape of rods or spatulas (Figure 153). They are often found associated with mosses. The few currently described species of the Xylonomycetes class live as endophytes of rubber trees. Only the anamorphs, which produce conidia protected in pycnidia, are currently known of these fungi. The line of Laboulbeniomycetes contains species whose biology is very particular compared to that of other Pezizomycotina. Indeed, they are epibionts of insects and have a defined development giving rise to a thallus with a constant number of cells. This is made up of a few tens of cells at most (often less) and measures a few tens of micrometers. The thallus is attached to the cuticle of an insect, often in a specific location, and it is not known if it causes discomfort for the insect (Figure 154). This thallus differentiates a small perithecium giving rise to ascospores which are sticky and ensure the spread of the fungus during contact between insects. Most species are homothallic, but there are a few heterothallics. There are several hundred species.

Figure 153.
Geoglossum fallax.
Figure 154.
Hesperomyces sp. on a ladybug.Leotiomycetes
Importantly, the class of Leotiomycetes contains several thousand species, including saprotrophs, mycorrhiza-forming species, necrotrophs and obligate parasites of plants. The fruiting bodies are often apothecia-like and very colorful (Figure 155), but Erysiphales are different by having characteristic cleistothecia (Figure 146, Erysiphe flexuosa [Translator’s note: This must point towards the wrong figure…]). Important pathogens include Sclerotinia sclerotiorum, Botrytis cinerea and Monilia (Figure 156). These fungi are necrotrophic pathogens with a broad host spectrum; e.g. up to 200 different species are attacked by Sclerotinia sclerotiorum. Botrytis cinerea causes “gray rot”. However, it is also used during the vinification of sweet wines such as Sauternes. Indeed, in certain climatic conditions, it causes a “noble rot” which allows the wine to acquire its richness in sugar and its characteristic taste. Monilia attack fruits such as apples, pears or quinces and cause brown rot, a dreaded disease by fruit growers.

Figure 155.
Two typical Leotiomycetes. Above, Bisporella citrina and Chlorociboria aeruginascens below.
Figure 156.
Monilia sp. on a quince.Sordariomycetes
The fifth and final class of Sordariomyceta is that of Sordariomycetes. This line contains more than 10,000 species that live either as saprotrophs or as parasites or mutualists of plants, animals or other fungi. Their fruiting bodies are perithecia that develop only after fertilization of the female gamete by a male gamete (Figure 146, Podospora anserina). These may not have an ostiole, but in this case a differentiated hymenium is still present, or be grouped in a large stroma as in Xylaria (Figure 157). It is in this group that we find the laboratory model fungi that are widely used for genetic and molecular analyzes: Sordaria, Neurospora and Podospora. These saprotrophs have rapid growth, simple nutritional demands and a short and perfectly controlled sexual reproduction cycle. They therefore serve as models for molecular genetic studies to elucidate multiple biological phenomena. Many species in this group are plant pathogens that pose big problems in agriculture. Magnaporthe oryzae is the main pathogen of rice, the most widely consumed cereal in the world. Fusarium graminearum (= Gibberella zea) is an important wheat pest that can pollute with mycotoxins. For the former, the infection is spread either by conidia or via the mycelium which then enters the plant through the root. For the latter, the infection is spread by ascospores. Claviceps purpurea which infects rye, is another famous species because of the alkaloids it produces, which if consumed, by eating bread made from contaminated flour, leads to ergotism associated with gangrene, loss of limbs and death. The alkaloids produced by this fungus cause vasoconstrictions. Numerous “epidemics” of ergotism have occurred in the past without, at the time, a cause being identified. Now these alkaloids are used in medicine but also as drugs, the famous LSD is a derivative of the alkaloids of this fungus! Tolypocladium inflatum, an endophytic species close to Claviceps, produces cyclosporin A which allowed the development of organ transplants because it is an immunosuppressant which has no side effects. Ophiostoma ulmi and novo-ulmi have destroyed elms in Europe and now in North America. Cryphonectria parasitica destroys chestnut trees in North America. The list of these important pathogens for human societies is still long! The pathogenic Sordariomycetes of animals are grouped mainly in the Cordycipitaceae family. Little is known about their impact on insect populations, but these fungi are collected very frequently, suggesting a major role in population control (Figure 158). Finally, there are some species which parasitize other fungi, such as Trichoderma (Figure 159). Trichoderma reseei, the only species of the genus which is saprotrophic, is widely used in industry as a source of various lytic enzymes (glucanases, cellulases, hemicellulase, etc.) which it secretes in the medium in large quantities.

Figure 157.
Carpophore of Xylaria polymorpha. On the right, the section shows the perithecia on the periphery embedded in the stroma.
Figure 158.
Cordyceps sp. on a caterpillar.
Figure 159.
Trichoderma sp. on a carpophore.Dothideomyceta
The second large lineage of Pezizomycotina is that of Dothideomyceta which includes six classes. Lecanoromycetes, Arthoniomycetes, Lichinomycetes and Coniocybomycetes group together lichens. All of them make apothecia, though these structures differ from class to class. Of these four classes, the most important is that of Lecanoromycetes since it includes 90% of known lichen species (Figure 160). A few species in this class are saprotrophs which appear to have lost the ability to live as a lichen. One species, Stictis / Conotrema, can also adopt both lifestyles. The class of Dothideomycetes contains more than 10,000 species presenting all the styles of life known to Eumycota: saprotrophs, lichens, parasites and mutualists of plants, animals etc. It is probably the most diverse class of Pezizomycotina. In this group, the fruiting bodies are pseudothecia (Figure 161) containing asci with a particular spore release mechanism that distinguishes them from other Pezizomycotina producing pseudothecia. The so-called bitunicated asci have in fact two walls which break sequentially. Important species include Alternaria which produce allergenic spores; Cochliobolus, Leptosphaeria, Pyrenophora and Mycosphaerella which infect many cultivated plants: rapeseed, wheat, corn, etc. Finally, among the saprotrophs, let us mention Baudoinia compniacensis which feeds on alcohol vapor and which can be found in whiskey distilleries or on Cognac rooftops!

Figure 160.
Three Lecanoromycetes lichens. From left to right: Xanthoria parietina, Peltigera horizontalis and Cladonia coccifera.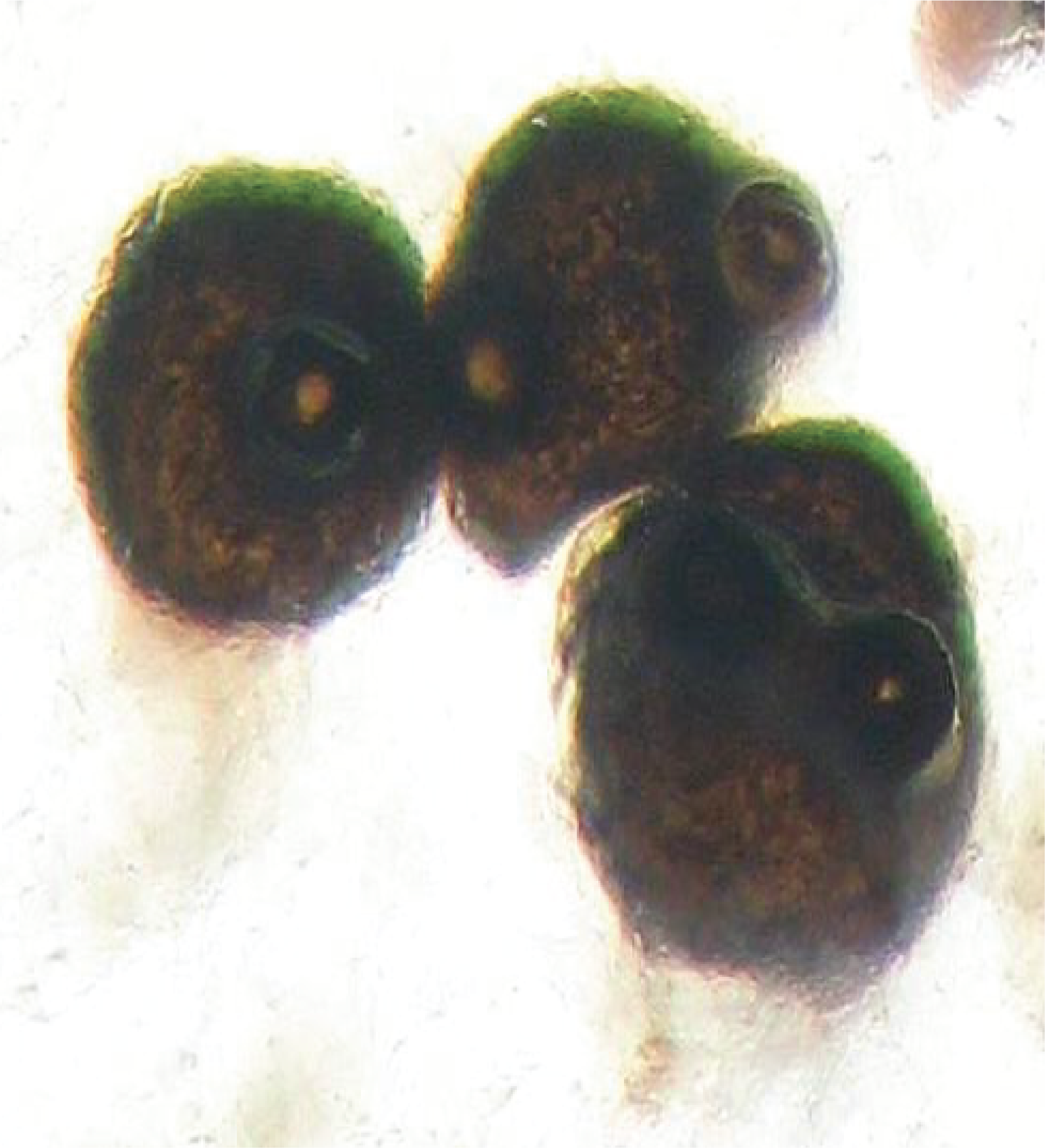
Figure 161.
Pseudothecia of Leptosphaeria maculans.Eurotiomycetes
The last class of Dothideomyceta and more generally of Pezizomycotina, that of Eurotiomycetes, which is probably the most important of all of them for human societies. In fact, this class includes important human pathogens, but also fungi used in industry and especially the fungus that led to the discovery of antibiotics, Penicillium notatum. There are several thousand species in this class. In nature, Eurotiomycetes are saprotrophs, which can sometimes attack plants, or lichens. The phylogeny strongly suggests that saprotrophic forms derive from lichens. These fungi have simpler fruiting bodies than those of other Pezizomycotina. They are cleistothecia or gymnothecia. The best known members are Aspergillus and Penicillium. The difference between the two genera is based on the shape of the conidiophores, but this character imperfectly reflects their phylogeny. Teleomorphs are often unknown because most species have no known sexuality. Perhaps to compensate, they produce incredible quantities of very volatile conidia. In fact, the majority of contaminations observed on food or materials are caused by Penicillium or Aspergillus (Figure 162). Aspergillus are used as study models in many laboratories but they are also widely used in the pharmaceutical and food industry. Aspergillus oryzae is used for making sake. It starts the degradation of rice starch and the sugars produced serve as the basis for fermentation by yeasts or bacteria. Aspergillus niger is used for the production of citric acid of which 500,000 tonnes are produced each year. Several species of Aspergillus produce mycotoxins (ochratoxins, sterimagocystins …) which are very toxic. As these fungi have the ability to grow in fairly dry environments, they can invade foodstuffs if they are not properly stored. The accumulation of mycotoxins can then become dangerous and lead to the loss of stored products. Penicillium are most famous for the production of antibiotics. In fact, the first antibiotic discovered and exploited was penicillin, first produced by Penicilium notatum, now by Penicillium chrysogenum (Figure 163). Nowadays, many antibiotics are chemically synthesized but some continue to be purified from fungi such as beta-lactams including penicillin G and cephalosporin C. It is in this genus that the famous Penicillium camemberti and Penicillium roqueforti are found, whose evocative names indeed indicate that they are involved in the manufacture of cheese (Figure 108).

Figure 162.
Aspergillus glaucus contaminating a jam jar.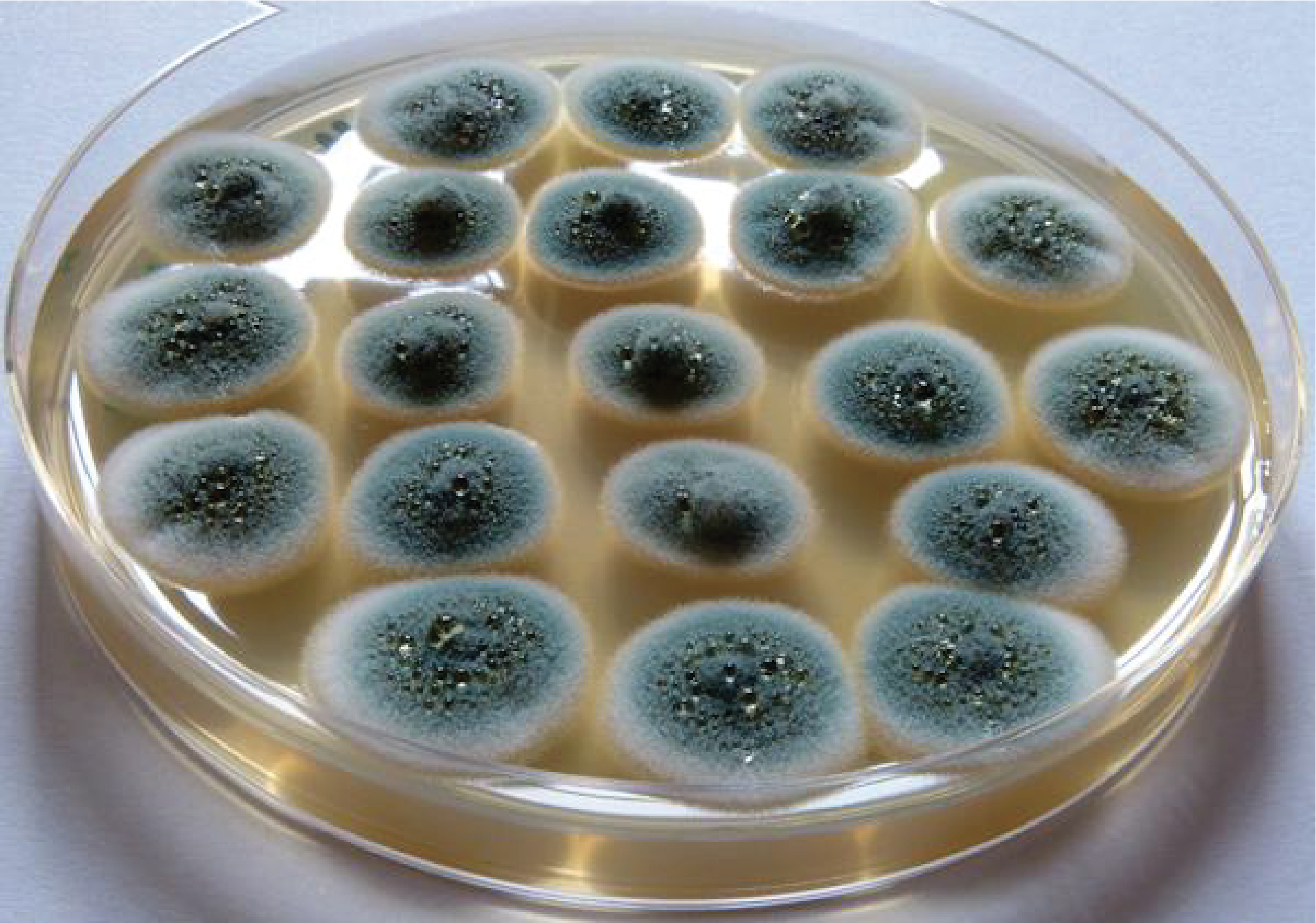
Figure 163.
Penicillium chrysogenum.Many Eurotiomycetes are important for public health. The most important in France is Aspergillus fumigatus which causes often fatal aspergillosis. It infects immunocompromised people, especially patients undergoing cancer treatment. Its spores are allergenic and small in size. They enter the body through the respiratory tract. The fungus therefore mainly causes pneumonia. Other important species belong to the order Onygenales, whose members are predominantly common soil saprotrophs. This order nevertheless contains several species which are true virulent pathogens because they are capable of infecting healthy subjects. However, they are also found naturally as saprotrophs in soils, showing that their parasitism is occasional and probably results from adaptation to their environment, rather than a strategy specific to parasitism. Contamination is also mainly through inhalation of spores and becomes severe rather in immunocompromised patients. The fungus undergoes a dimorphic transition to a yeast form in the human body, as it grows as a filament in nature. Finally, there are many species capable of persisting in the skin or nails where they cause athlete’s foot, ringworm or onychomycosis.
Back to chapter index