Basidiomycota
Back to main indexJump to section:
Introduction
The branch of Basidiomycota, or Basidiomycetes, forms the second large phylum of Eumycota since it contains 30% of the species of described fungi. A shared trait of all Basidiomycota is the production of their meiospores by budding of specialized cells, the basidia. The spores are therefore called basidiospores (Figure 164). In addition, the majority of species has a dikaryotic mycelium originated from plasmogamy of sexually compatible cells. In the Ascomycota, only Taphrinomycetes presents such a mycelium, as the dikaryotic stage is non-existent among Saccharomycotina and restricted to within the fruiting body in Pezizomycotina.
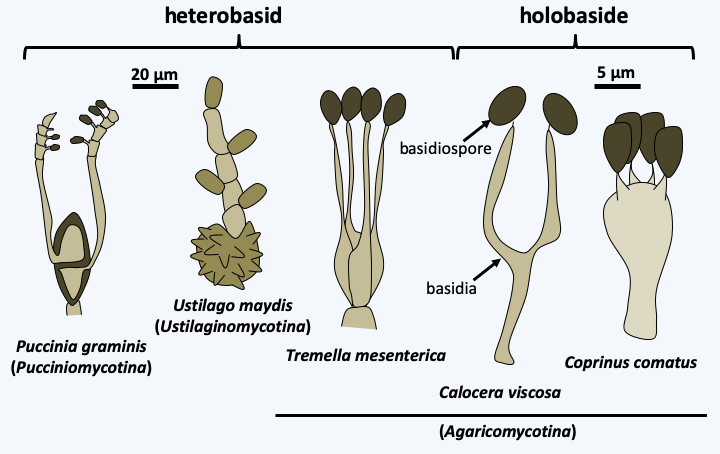
Figure 164.
Basidia and basidiospores.The dikaryotic state is maintained according to the scheme shown in Figure 131. This mechanism produces a mycelium bearing a characteristic structure at the septa between the cells called a clamp connection (Figure 165). The observation of clamp connections is therefore a good diagnostic character to determine if a mycelium comes from a Basidiomycota, in addition to being dikaryotic. Nevertheless, all Basidiomycota do not produce clamp connections even at the dikaryotic stage, and absence of clamps is therefore not a good criterion for excluding such species from the phylum. Molecular phylogenies largely confirm the monophyly of the Basidiomycota and define three sub-branches: Pucciniomycotina, Ustilaginomycotina and Agaricomycotina (Figure 166).
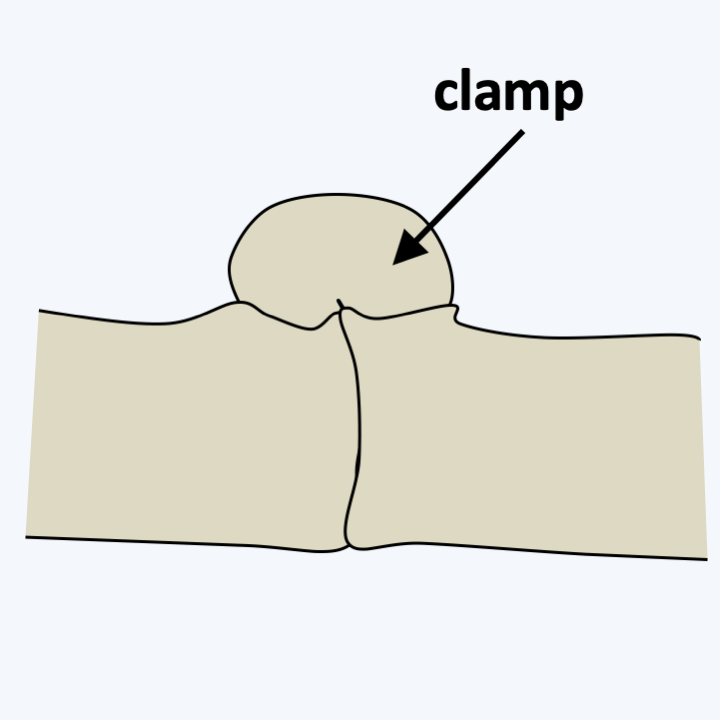
Figure 165.
Anastomosis clamp.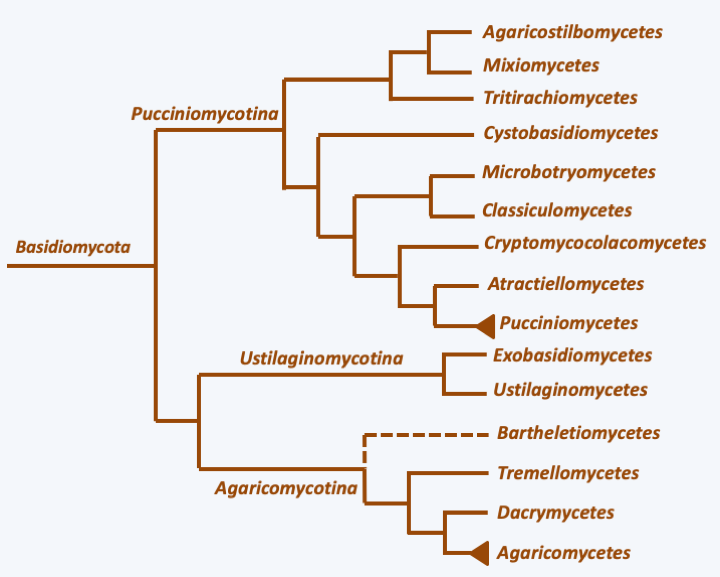
Figure 166.
Phylogeny of Basidiomycota.Pucciniomycotina
The Pucciniomycotina, formerly Urediniomycetes, include ~7,500 described species. They are mostly obligate parasites of plants, especially gymnosperms and angiosperms, which appears to be the ancestral lifestyle of species in the group. Others live as saprotrophs or parasites of insects and mushrooms. Some species have adapted to aquatic life and are found in fresh or marine waters. Phytopathogenic species are often obligate biotrophs and can have life cycles that require two hosts and involve up to 5 different dispersal propagules! In many species the spores are numerous and reddish, hence the name “rust” given to these fungi. Some rusts cause considerable damage to crops, especially cereals. The septal pores of rust fungi are simple and sometimes have corpuscles resembling Woronin grains typical of Pezizomycotina (Figure 167). Few species form clamp connections in the mycelium. Some species produce simple, often small, multicellular fruiting bodies. Others have regressed to forms dividing essentially like yeasts. These often live in association with plants and in particular form the majority of the biomass of the phylloplane. Of the nine classes recognized in the subphylum, seven contain only a few described species. Of the last two classes, that of Pucciniomycetes is the most species rich and the most important in ecosystems and agriculture, while that of Microbotryomycetes is known from evolutionary biology studies which are carried out using some of its representatives, e.g. Microbotryum violaceum.
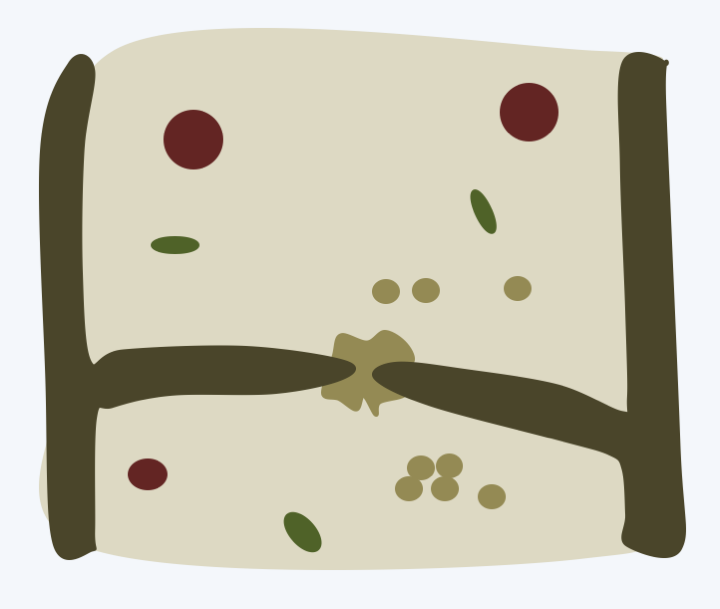
Figure 167.
Simple pore of Pucciniomycotina.Pucciniomycetes
The class of Pucciniomycetes, and more particularly the order of Uredinales, encompasses 90% of the described species of Pucciniomycotina. They are obligate biotrophic plant pathogens attacking many hosts, ranging from trees such as pines to grasses and shrubs such as raspberries (Figure 168). The best known species is Puccinia graminis which infects grains and causes black rust in wheat. Other wheat rusts are caused by Puccinia striiformis for yellow rust and Puccinia recondita for brown rust. In antiquity, these species caused such devastation that the Latins at the beginning of spring celebrated the “Robigalia”, a festival to appease the god Robigus who was supposed to be responsible for the devastation to crops caused by Puccinia graminis. The life cycle of Puccinia graminis is the most complex among eukaryotes because it involves two hosts and five forms of spores (Figure 169). Another order of Pucciniomycetes, comprising about 170 species, is that of the Septobasidiales. This is a group of parasites and mutualists of scale insects such as mealybugs. Indeed, while some species clearly behave like parasites of these insects, others form a structure that in its functioning resembles a lichen, except that the phycobiont is held by the insect! The life cycle of Septobasidiales begins like Puccinia graminis with a septate basidium that releases its basidiospores. These can germinate into yeasts which generate colonies when cultured. When the spores/yeast come into contact with a mealybug, they generate a mycelium which will proliferate to form a thallus above a leaf or branch harboring mealybugs (Figure 170). The thallus contains chambers which house the animals. Parts of the mealybugs are penetrated by hyphae of the fungus. These insect individuals are sterile, smaller than the rest and bring food to the symbiosis partners by sucking the sap of the plant. The other mealybugs are free, fertile and… protected by the mycelial thallus of the fungus! On the upper surface of the thallus the basidia are produced. The multicellular thallus thus also serves for fruiting. These associations can have a strong impact in agriculture. Indeed, Septobasidium pseudopedicellatum is an important pathogen of lemon trees in the southern United States, Septobasidium bogoriense attacks to coffee and Septobasidium theae is a pathogen of tea plants.

Figure 168.
Phragmidium rubiidaei, raspberry rust.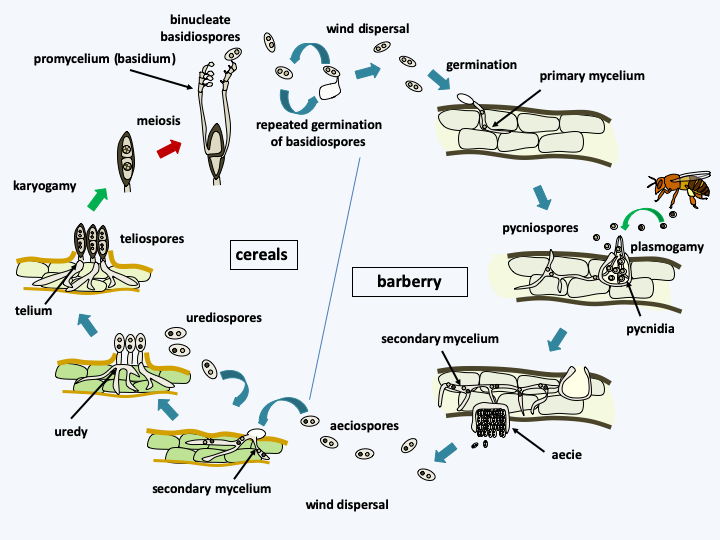
Figure 169.
Puccinia graminis life cycle. The cycle begins in the spring with the germination of bicellular and diploid teliospores, followed quickly by meiosis and the formation of basidiospores on a heterobasid or promycelium. Basidiospores are actively released and can germinate repeatedly. They are dispersed by the wind and cannot infect cereals. They arrive on the secondary host, barberry Berberis vulgaris, in which a haploid and monocaryotic mycelium or primary mycelium is formed. Pustules, actually pycnidia, form on the upper surface of the leaves of the plant. They contain pycniospores which will serve as male gametes and receptive hyphae which will serve as female gametes. A sweet solution attracts insects which will ensure the propagation of the pycniospores to strains of compatible sexual types. When a pycniospore arrives on a receptive hyphae of compatible sexual type, cell fusion occurs and gives rise to the dikaryotic mycelium called secondary mycelium. It produces aeciospores in aecia which form on the underside of the leaves. Aeciospores cannot re-infect barberry. They are dispersed by the wind and go to contaminate cereals, where it gives rise to a dikaryotic secondary mycelium. This mycelium differentiates from uredias containing urediospores which will amplify the infection on cereals until autumn arrives and the formation of bicellular teliospores occurs, first dikaryotic, then diploid. These are very resistant and will spend the winter in rest. Knowing the cycle has drastically reduced the problems caused by this rust because it was enough to remove its alternative host, barberry, to decrease the infection.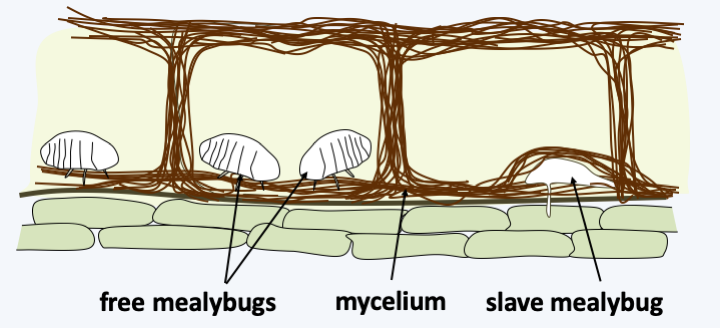
Figure 170.
Septobasidium theae and its mealybugs.Microbotryomycetes
The class of Microbotryomycetes contains saprotrophic and parasitic species, notably parasites of plants such as Microbotryales. These parasites have a life cycle resembling that of the smuts (Ustilaginomycotina) with which they have long been classified (Figure 171). Microbotryum violaceum, the model species of the group, is able to infect a hundred Caryophyllaceae species where it attacks the anthers, resulting in a sexually transmitted disease (Figure 172). This disease is very common and infected catchflies, campions or soapworts can be frequently encountered in nature. Fungi of the Microbotryales are used as models in genetics to understand the dynamics and evolution of fungal infections in plants, in addition to different modes of sex chromosome evolution.
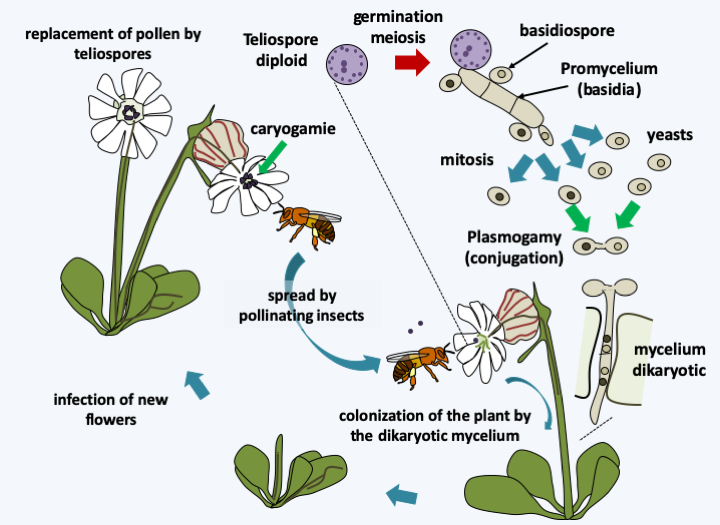
Figure 171.
Cycle of Microbotryum violaceum. Diploid teliospores are carried from flower to flower by insects. They germinate to give rise to a promycelium carrying basidiospores. Their germination gives rise to haploid cells which spread like yeast. The fusion of two sexually compatible yeasts generates a dikaryotic mycelium which infects the host plant to return the teliospores to the level of the anthers of the new flowers, where they await the insect which will disperse them.
Figure 172.
Microbotryum violaceum on soapwort. The purple spores of the fungus have replaced the normally translucent pollen grains on the stamens.Ustilaginomycotina
The subphylum Ustilaginomycotina includes about 1700 species, most of which are parasites of flowering plants. Most species infect a single host. Others, like Malassezia, colonize the skin of mammals, including humans, or feed on secretions from the sebaceous glands at hair follicles. Phytopathogenic species attack flowers and prevent seed formation, thereby having an important economic impact. Such species often alternate between an infectious mycelial phase and a saprotrophic yeast phase. They attack their host in a special way by secreting deposits inside plant cells via characteristic vesicles. At the end of infection, they often form clumps of black spores, giving them the common name “smut fungi”. Their septal pores are more complex than in Pucciniomycotina (Figure 173). In several orders, this pore is not present and has probably disappeared by regressive evolution. Two classes have been defined on the basis of molecular data.
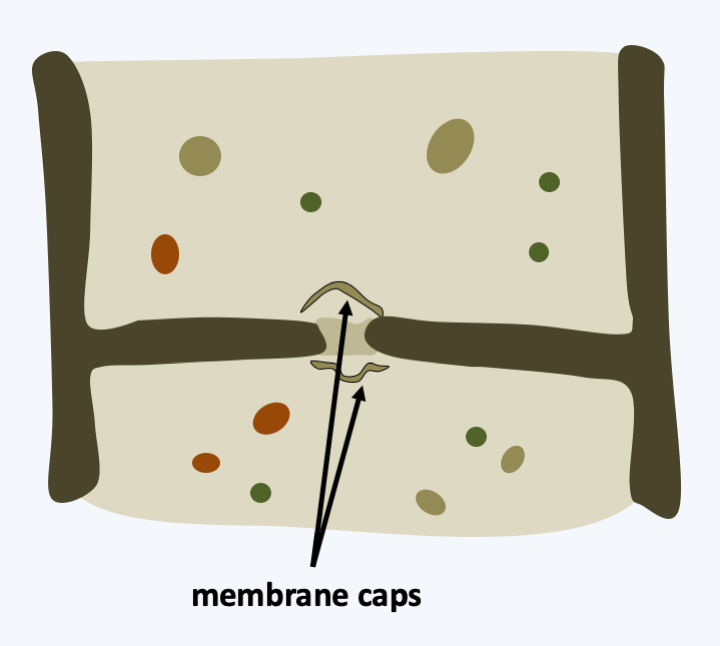
Figure 173.
Septal pore in Ustilaginomycotina. The pore is surrounded by vesicles deriving from the reticulum called membrane caps.Ustilaginomycetes
The Ustilaginomycetes class contains the best-known species of the subphylum, Ustilago maydis, which attacks maize and generates galls. The galls are effectively tumors where the host’s tissue is enlarged and mixed with the mycelium, replacing kernels in the cob (Figure 174). The life cycle is very similar to that of Microbotryum violaceum, but the dispersal of teliospores is by wind and not by insects. In addition to its destructive aspect of crops, certain species or strains of Ustilago can cause serious damage to human health. Consumption of parasitized corn by Ustilago maydis is widespread in Mexico where galls are highly prized and sold under the name of Huitlacoche (Figure 175). Their ingestion may cause various ailments known as ustilaginism, especially in children. The fungus produces various toxins, alkaloids among others, some of which inhibit the action of adrenaline. It also causes abortions in cattle. The rest of the class have similar life cycles and infect many plants.
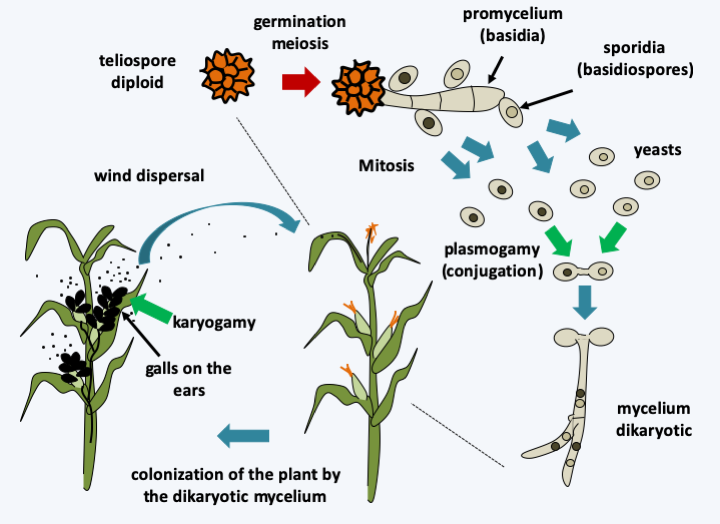
Figure 174.
Ustilago maydis life cycle. The cycle begins in the spring with the germination of the diploid teliospore accompanied by meiosis to give rise to a promycelium of 4 haploid basidia. Each of these cells produces at least one bud, the sporidia, which is the equivalent of a basidiospore. Budding can be repeated several times and therefore produce yeast colonies. If sexually compatible cells meet, cell fusion occurs and the dikaryon is formed. This leads to the formation of the curly mycelium and the invasion of the plant, which therefore only occurs if there is sexual fusion. When it reaches the surface of the plant, the mycelium can also sometimes produce dikaryotic spores which ensure asexual spread. Nuclear fusion occurs in the teliospores that form during the fall in galls which occupy the place of grains in the ear, terminating the cycle and ensuring the production of a form of resistance during the winter.
Figure 175.
Packages of Ustilago maydis or huitlacoche (center) sold among other vegetables and mushrooms at a market in Mexico.Exobasidiomycetes
The class of Exobasidiomycetes also contains important pathogens, including Tilletia caries which is a pathogen of wheat, causing decay. Its life cycle resembles that of Ustilago maydis. Young plants are infected in the spring and the infection is asymptomatic until harvest, where the grain is replaced by the powdery mass of spores. The spores have a longevity of at least 15 years in the soil. The life cycle of this fungus was determined by Isaac-Bénédict Prévost (1755-1819) at the beginning of the 19th century, who thereby showed for the first time, long before Robert Koch, that the cause of a disease was in fact a microbe. The Malassezia are other important members of the class. These commensal yeasts in the skin can under certain ill-defined conditions proliferate and cause dermatitis including scalp dandruff or tinea versicolor.
Agaricomycotina
The subphylum of Agaricomycotina, formerly Hymenomycetes, can be considered as the counterpart of Pezizomycotina for the Basidiomycota, because it contains the majority of the described species, i.e. more than 20,000. It encompasses the majority of familiar fungi that produce large carpophores, which were used to establish the first classifications (Box 18). Some are edible such as button mushrooms, chanterelles, boletes, sheep’s foot etc.; others are toxic such as fly agarics, galerinas etc. Some species with very similar appearances can be some edible and others very toxic or even fatal. The number of “specialists” having passed to the other side because of mushrooms is not counted any more. I urge you for more information to go see “Le Roman d’un Tricheur” by Sacha Guitry or listen to Mireille, Pills and Tabet in “Flowers and Mushrooms”.
Box 18. Fruiting body morphology in Agaricomycotina

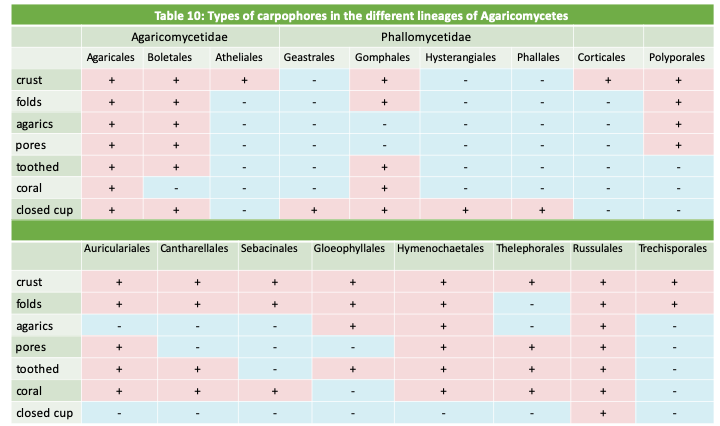
As in the Ascomycota, the shape of the fruiting body used to be an important feature in the classification of Agaricomycotina. Molecular phylogenies have revealed many instances of convergent evolution with the repeated occurrence of most forms, especially in the class of Agaricomycetes (see Table 10). In the old classification, the part of the Agaricomycotina producing septate basidia, now the class Tremellomycetes and the order Auriculariales, were grouped together with Ustilaginomycotina and Pucciniomycotina in the now defunct class of Phragmobasidiomycetes. Species differentiating non-septate basidia were placed in the class of Holobasidiomycetes, itself separated into two subclasses: Hymenomycetes with a hymenium exposed to the open air and Gasteromycetes with an unexposed hymenium. Note that these groups nowadays do not reflect phylogenetic relationsips but are still sometimes used for descriptive purposes.

Among the Hymenomycetes which produce the classic fungus with a central foot (or stipe) and a cap (or pileus), mycologists distinguished the 'agarics' with the hymenium positioned on lamellae, the 'boletes' where the hymenium lines the insides of pores and 'hydnoids' where it is on prickles. Other forms frequently encountered are the 'crusts' and 'jellies', the 'clavaries' in the form of corals and the 'polypores' in the form of consoles.

Gasteromycetes exhibit a wide variety of forms. The most famous are the puffballs. Other forms resemble stars or bird nests. Without question, the strangest carpophores are found in the order of the Phallales. They take on shapes ranging from a brightly red-colored star, to cages, flowers or phalluses!

The purpose of these different carpophores is to efficiently disperse the spores. For the Hymenomycetes, where the main vector is the wind, the main evolutionary tendency is to ensure a departure of the spores from as high up as possible, hence the repeated evolution of the foot; the ancestral form being the crust type. The purpose of the lamellae, pores and prickles is to increase the spore production area. In Gasteromycetes, rain and/or animals play a major role. For example, the fructification of bird's nest fungi is studied for the dispersal by drops of rain, which when they splash the inside of the cup, propel the 'peridioles', the small, egg-like masses containing the spores. That of the Phallales are often brilliantly colored, even like Aseroe rubra resembling a flower, because it performs the same function: to attract insects. Here, the target audiences are flies and therefore the smell of this type of fruiting body mimics that of animal corpses. The attracted flies consume the gleba, the blackish substance containing the spores. Agaricomycotina have therefore been no less inventive than plants and animals when it comes to evolving strategies to disperse!
Agaricomycotina lifestyles are very varied. The majority of the species are saprotrophs involved in the recycling of wood and other plant materials. Some are very effective for the degradation of cellulose and lignin because they produce and export numerous enzymes (peroxidases, laccases, hydrolases, etc.) to the extracellular environment. A few species are lichens and many others live as mycorrhizal mutualists. Several Agaricomycotina are able to attack healthy or weak plants, in particular trees, and are therefore dangerous pests. Others are mycoparasites or animal parasites. One genus contains important human pathogens causing systemic mycosis mainly in immunocompromised patients. The septal pores are called “dolipores”. They have a typical morphology with associated vesicles which derive from the endoplasmic reticulum (Figure 176). In section, they appear as a dotted parenthesis, hence their name parenthesome. In reality, it is a continuous cuff pierced with small pores. The number of pores varies in different classes. They are absent in species that diverged the earliest from the rest, e.g., Tremellomycetes, and numerous in other Agaricomycetes. The parenthesome contains characteristic proteins present only in Agaricomycotina. The dolipores are necessary for the proper functioning of the mycelium and for the development of fruiting. Indeed, the deletion of a gene encoding the spc33 protein in Schizophyllum commune leads to a decrease in growth rate and a reduction in the production of carpophores. The mycelium of this mutant does not close its septal pores upon injury. The mycelium of Agaricomycotina, provided it contains functional dolipores, is capable, like that of Pezizomycotina, of supporting the production of complex multicellular fruiting bodies. However, unlike Pezizomycotina, dispersal by asexual spores appears to be less widespread. The generic life cycle of these fungi is presented inFigure 177. However, large variations exist around this cycle with different modalities for the production of basidia and basidiospores and especially for the development and morphology of the fruiting body (basidiocarp). The fruiting bodies can have the classic appearance of a “mushroom” but also appear as crusts, jellies, brackets, corals, wineskins, stars, phalluses or even bird nests! These forms are related to the mode of dispersion (Box 18).
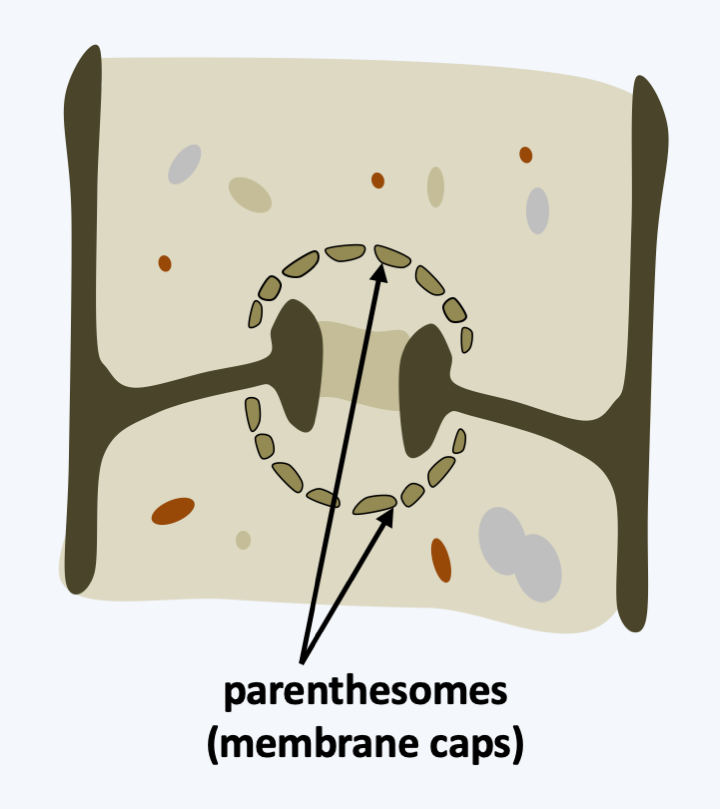
Figure 176.
Dolipore (septal pore) of Agaricomycotina.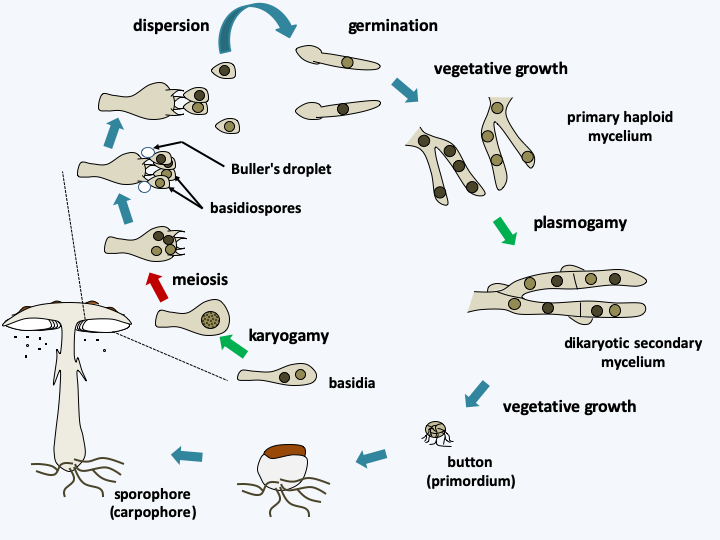
Figure 177.
Generic life cycle of Agaricomycotina. The cycle begins with the germination of basidiospores which give rise to so-called primary haploid mycelia. These are generally not very combative and have reduced capacities to proliferate. The meeting of two primary mycelia of compatible mating types results in the formation of the dikaryotic mycelium with clamp connections. This has much greater growth potential. When conditions are right, often when the mycelium is deficient, pimples are differentiated. Some of them will develop into carpophores carrying basidia, then basidiospores. In the Hymenomycetes which disperse via the wind, the spores carried by the sterigmas, which are the small growths at the top of the basidium, are expelled by an original mechanism. A sphere of liquid appears at the base of the basidiospore, the Buller's droplet, on a special structure of the spore called the hilar appendage. The mechanism involved is still debated. This drop coalesces with another liquid layer affixed to the surface of the spore, causing a change in the center of gravity of the whole. This leads to the detachment of the basidiospore from its sterigma with an acceleration of the order of 10,000g. The distance traveled is sufficient to move far enough away from the basidium without meeting the wall of the hymenium located opposite! The spore then falls by gravity and is then dispersed by the wind. The manufacture of gout explains the need for humidity in the air for the dispersion of these fungi. This mechanism also explains the general morphology of Agaricomycotina fruiting bodies: a foot to raise the hymenium, the inferior position of the fertile tissue so that the spores fall and the cap for its protection. The spacing of the blades, the diameter and the shape of the pores are also related to this expulsion mechanism. Other strategies are described in Box 18.Currently, the molecular phylogeny of Agaricomycotina defines four classes of varying importance. The first, that of the “Bartheletiomycetes”, does not yet have the formal rank of class and contains only one species, Bartheletia paradoxa (Figure 178). This fungus is a saprotroph that lives on the leaves of Gingko biloba. Gingkos are relics of a group of gymnosperms abundant in the secondary era. They are therefore considered “living fossils”. Interestingly, Bartheletia paradoxa presents, like its substrate, archaic characters, with the presence of micropores rather than a central pore in the septa between cells and “teliospores” which give rise to basidiospores, as in Pucciniomycotina and Ustilaginomycotina. It would thus seem that this species too is a “living fossil”. The molecular phylogenies confirm its position at the base of Agaricomycotina, but do not allow us to decide whether this species defines a new class, or whether it is included in that of Tremellomycetes.
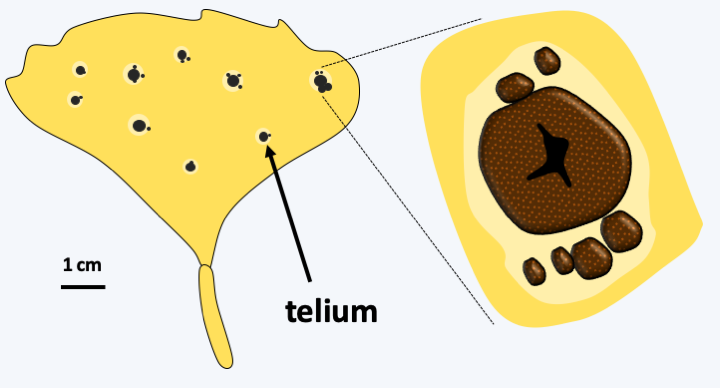
Figure 178.
Bartheletia paradoxa on a Gingko biloba leaf. Telia (sing. Telium) differentiate resistant diploid spores that can persist in the environment. Their germination gives rise to basidiospores.Tremellomycetes
The class of Tremellomycetes mainly includes saprotrophs and parasites of other fungi. These Agaricomycotina always present archaic characters. The septal pores have unperforated or perforated parenthesomes but with cup-shaped vesicles. Some species produce teliospores. The basidia are in most species transversely septate (Figure 164, Tremella mesenterica). The haploid stage is yeast-like, similarly as in some Pucciniomycotina and Ustilaginomycotina. The sexual fusion of two compatible yeasts gives rise to a dikaryotic mycelium that differentiates an often gelatinous carpophore (Figure 179). The life cycle therefore resembles that of Ustilaginomycotina. Other species live mainly in the form of yeasts. This is the case for two human pathogens, Cryptococcus neoformans and Cryptococcus gattii that develop as capsulated budding yeasts (Figure 180). They are found naturally in soils, especially those rich in pigeon droppings or on certain trees such as eucalyptus. Their sexual reproduction requires a large amount of inositol, so these fungi only do so in nature on eucalyptus trees which contain a lot of it. Teleomorphs, called Filobasidiella neoformans and Filobasidiella bacillispora, do not produce carpophores. These fungi are heterothallic with two mating types, MATa and MATα. When there is a nitrogen deficiency, two compatible cells merge. A curly dikaryotic hypha forms and gives rise at its tip to a holobasid, that is, an unseptate basidium. This basidium produces four basidiospores which immediately divide to give rise to yeast chains (Figure 180). In the laboratory, it is possible to induce a “monokaryotic” sexual cycle. In this case, the mycelium results from the fusion of two nuclei of the same mating type and gives diploid hyphae with incomplete clamp connections. Meiosis normally takes place in the basidium located at the end of the hyphae. In nature, only the mating type MATa is common. Cryptococci cause cryptococcosis. The infection is chronic and begins in the lungs after inhalation of infectious propagules. In immunocompetent subjects the lesions remain localized, but in immunocompromised patients, the fungus spreads with a predilection for the nervous system. Some antifungals, which lose their effect quickly because resistance develops during treatment, or the surgical removal of localized lesions can fight these parasites although they generally lead to death. Currently, there are 300 million cases annually, mainly in AIDS patients. Note that Cryptococcus gattii can infect non-immunocompromised individuals and cause serious damage. An epidemic involving hundreds of patients recently took place in Vancouver, Canada. Many eucalyptus trees had been planted in the area around the city… Cryptococcus are able to persist in the body to reappear later. In fact, they can persist inside macrophages for a long time. This is probably due to the fact that in their natural environment, they are the prey of amoeba. During evolution, they developed a strategy to escape their predator by inhibiting digestion by hydrolytic enzymes in the lysosome. The key elements of this escape are the establishment of a melanin coating and the presence of the glucuroxylomannan capsule. The same mechanisms are involved for persistence inside macrophages!

Figure 179.
Carpophore of Tremella foliacea.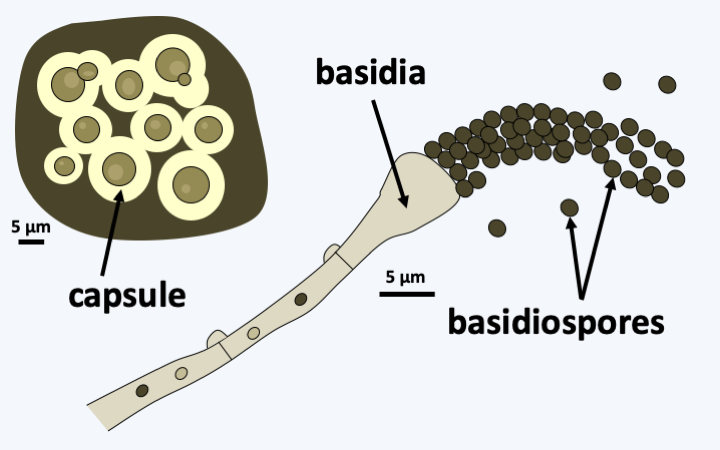
Figure 180.
Anamorph and teleomorph of Cryptococcus (Filobasidiella) neoformans.Dacrymycetes
The class of Dacrymycetes contains about 100 species. They are common saproptrophes. They are characterized by their dolipore having the parenthesome pierced by a single central pore, in addition to their fork-shaped basidium with two peaks (Figure 164, Calocera viscosa). Their fruiting bodies are often brightly colored orange / yellow and exhibit the shapes of brains, cupules or corals (Figure 181).

Figure 181.
Dacryopinax spathularia.Agaricomycetes
The last class, that of Agaricomycetes, is the numerically largest one since it includes more than 20,000 described species. Their life cycle, their perforated dolipore mycelium and their holobasids are typical of Agaricomycotina with a few exceptions such as certain Auriculariales, which is the group that diverged the earliest. Agaricomycetes are mainly saprotrophs, in fact the most efficient in the living world to degrade lignocellulose, and mycorrhizal mutualists. Some species are pathogens attacking trees, such as Armillaria, Heterobasidion annosum which parasitizes conifers, or Moniliophthora perniciosa which attacks cocoa trees, and some species are even lichens. Agaricomycetes are probably the most common fungi in the soil, although they are less often isolated in cultivation than Ascomycota or Mucoromycotina due to their slower growth than those of other Eumycota. They have diversified to an astonishing extent and the strategies they have developed to disperse are multiple, resulting in fruiting bodies of widely varying shapes, sizes and colors (the fungi pictured in Box 18 are all Agaricomycetes).
The details of the Agaricomycetes classification are complex and beyond the scope of this book. All the more so as their fruiting body morphologies were established via multiple evolutionary convergences, making it difficult to establish family links by simple analysis of the morphology of carpophores and spores. In addition, there are still many species that have not yet been analyzed and many lineages are yet to be discovered. Table 10 gives the main lines currently defined and the types of fruiting bodies they have established. It is easy to see from this table that the main types of carpophores have been invented by many lines! Detailed analysis of the results of the phylogenies indicates that the ancestral fruiting of the group would be of the “crust” type. Note that it is likely that species producing crusts that have yet to be analyzed, or even discovered, may be members of evolutionary lines additional to those indicated in the table, because such species have very similar morphologies and are often inconspicuous and therefore little studied.
Agaricomycetes are collectively very important in nature as they actively participate in soil formation. They ensure health because they manufacture humic acids which retain mineral salts; humic acids are degradation products of lignin. These fungi are also at the base of the food chain leading to small animals which participate in recycling plant waste. As mycorrhizals, they participate in the mineral nutrition of many plant species, especially trees in boreal regions. They also have an important role in the nutrition of “large animals”, such as squirrels, wild boars and… humans! In fact, the majority of “mushrooms” consumed (Figure 182) belong to this class, such as chanterelles, porcini mushrooms, trumpets of the dead, field mushrooms, button mushrooms, shiitake, oyster mushrooms, milk-caps, sheep’s feet, ox tongues, wood blewits, etc. Some species, such as ink caps or Schizophyllum commune, are laboratory models for studying the development of fruiting, in particular the meiosis stage, or the structure and function of the mating type (Figure 183). Others are analyzed for their content in genes which encode proteins capable of breaking down complex carbohydrates, because their enzymes have practical applications in paper bleaching or in the manufacture of biofuel from dead plant material.

Figure 182.
Some edible species of Agaricomycetes. From top left to bottom right: Chanterelle Cantharellus cibarius, Death Trumpet Craterellus cornupioides, Oyster Mushroom Pleurotus ostreatus, Milkweed Lactarius deliciosus, Mutton's Foot Hydnum repandum and Ox's Tongue Fistulina hepatica.
Figure 183.
Two laboratory models. The coprins of herbivorous droppings such as Coprinopsis cinera or Coprinopsis sp. in the photo on the left and Schizophyllum commune in the photo on the right are widely used as models in laboratories.Back to chapter index