Holozoa
Back to main indexJump to section:
Introduction
Recent phylogenies define the Holozoa as the line of Opisthokonta that includes animals (Metazoa). This lineage also includes a paraphyletic group of unicellular organisms possessing various types of biology, the Choanozoa (Figure 86); phagotrophs, parasites, unicellular, colonial… From an anthropomorphic point of view, this line is of the greatest importance because it gives us an idea of the aspect of our unicellular ancestor.
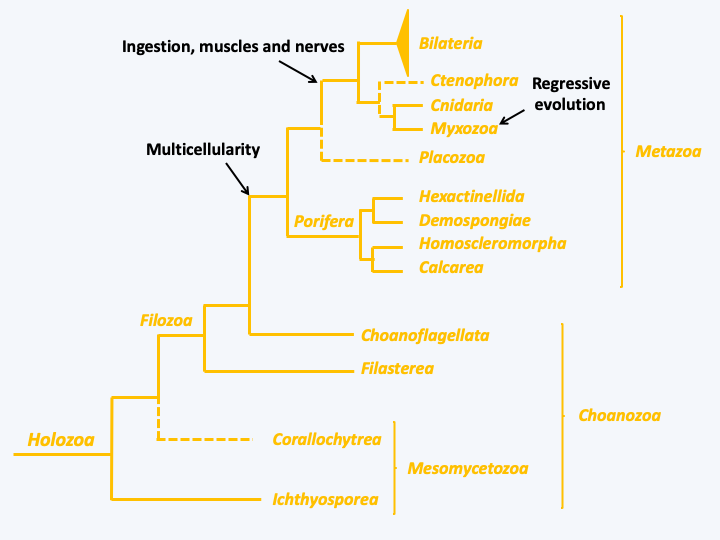
Figure 86.
Phylogenetic tree of Holozoa.Mesomycetozoa
At the base of this tree is the Mesomycetozoa, which means intermediary between fungi and animals. Indeed, most of the organisms in this group have been classified with the fungi in the old extinct class of Trichomycetes, because they have fungal characteristics such as the possession of a vegetative thallus formed of hyphae and the production of spores to disperse (Figure 87). The forty or so known species are commensal or parasitic symbionts of fish, amphibians or crustaceans. Nonetheless, metagenomics shows that these organisms are frequent and diverse in marine and freshwater ecosystems, indicating that only a small proportion of them are known to science. Few species are able to grow in vitro, and it is likely that the complete life cycles are not known (Figure 87). The ones that are possible to cultivate grow in axenic media, a characteristic which rather brings them closer to fungi. Creolimax fragrantissima can be transformed and analyzed genetically by an RNA interference approach, so this species will likely serve as an analytical model for Mesomycetozoa. Another species, Corallochytrium limascisporum, is related to this group, or may even belong to it. It would then form the second class of the group, the Corallochytrea, the other species being currently included in the Ichthyosporea. It has a similar life cycle involving amoeba for dispersal and isolated cells dividing by fission during the trophic phase. In the wild, it lives in the coral reefs of the Arabian Sea. It is very easily cultivated on an axenic medium and does not require organic nitrogen to grow, a character which makes it similar to fungi.
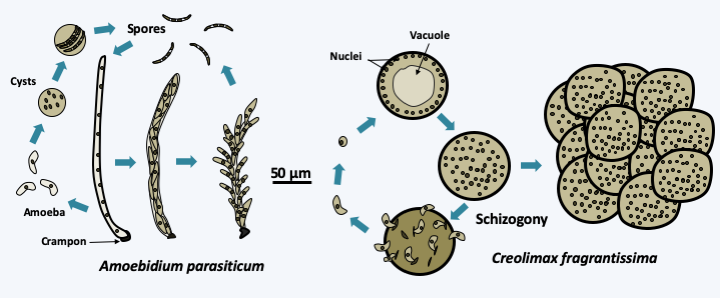
Figure 87.
Asexual cycle of two Mesomycetozoa. Amoebidium parasiticum has long been classified with fungi because it differentiates an elongated, hypha-shaped vegetative thallus and disperses via spores. This species is found attached by a spike to the outer cuticle of crustaceans and freshwater insect larvae. It can differentiate between amoebae which will emerge from the thallus and encyst and give back spores. This part of the cycle is similar to that of Creolimax fragantissima. This symbiont of the digestive tract of marine invertebrates produces amoebae which will grow in size and form syncytia. New amoebae emerge after syncytium schizogony. As cultures age, syncytia remain clumped together and form a structure that resembles a morula.Filastera
The Filastera class currently contains only two species: Ministeria vibrans and Capsaspora owczarzaki (Figure 88). Ministeria vibrans is a marine phagotroph that feeds on bacteria. It is a small radiant cell that lives either in free form or attached to the substrate by a long stem. Little is known about this organism. In contrast, a lot of data is available on Capsaspora owczarzaki, which is a symbiote of an aquatic gastropod, Biomphalaria glabrata. This mollusk is a vector of the flatworm Schizostoma mansoni, a human parasite that causes intestinal schistosomiasis. Capsaspora owczarzaki consumes the larvae of the worm and is therefore considered as a control agent of the parasite. It occurs as a radiant amoeba which can differentiate into a cyst or aggregate randomly. The genome of this organism has been sequenced and it contains around 8,700 genes for a size of 28 Mb. Among these, genes involved in multicellularity in animals have been found, such as extracellular matrix proteins, adhesins or tyrosine kinases. The expression of the genes encoding these proteins is induced during aggregation, which is remarkably similar to what happens in animals!
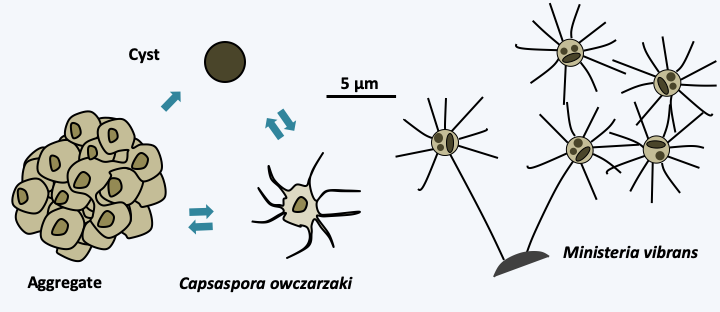
Figure 88.
Capsaspora owczarzaki and Ministeria vibrans, the two known Filastera.Choanoflagellata
The last class of unicellular Holozoa is that of Choanoflagellata. It is by far the best known, although there are still many gaps in our knowledge, and the one that contains the most species, since around 150 morphospecies have been described. This number probably underestimates the diversity of the group because some morphospecies, such as Codosiga botrytis (Figure 89) are so genetically variable, that new genera are likely to be created to classify the different lineages. Molecular phylogenies confirm that it is the group of organisms that is closest to metazoans. As the Choanoflagellata strongly resemble the choanocytes of Porifera, a close relationship between these groups has been considered for quite some time. Choanoflagellata inhabit fresh or marine waters. The typical cell is a small uninucleated cell characterized by the presence of a single flagellum surrounded by a collar (Figure 89). This collar is composed of microvilli agglomerated to each other. The beating of the flagellum attracts food particles like bacteria and small eukaryotes to the collar which carries them to the cell body, where they are ingested by phagocytosis. This mode of nutrition seems effective because from an ecological point of view, Choanoflagellata can represent up to 40% of small coastal flagellates and filter between 10 and 25% of coastal waters. They thus eliminate many bacteria by feeding on them. The group also participates in the elimination of viruses, because it has been shown that in their presence, viral particles are ingested and eliminated. They themselves serve as food for many organisms, e.g. Ciliophora…
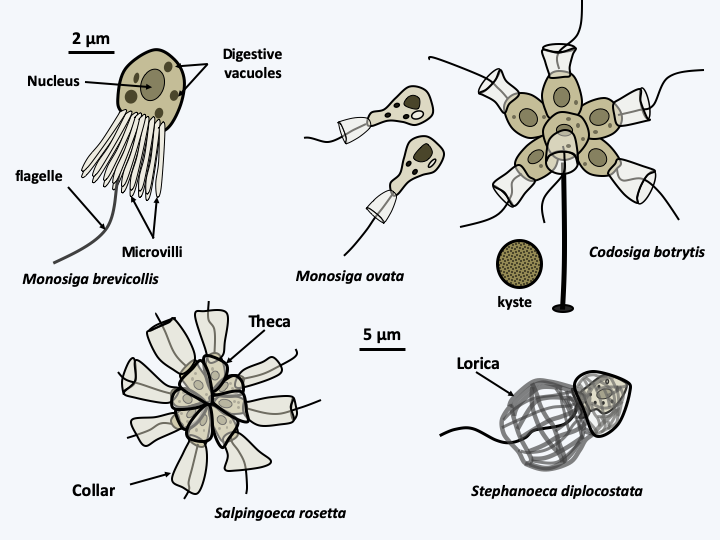
Figure 89.
Diversity of Choanoflagellates. Top left, the typical cell with its frill of microvilli surrounding the single central flagellum. The cell body contains a central nucleus. Prey are attracted to the water current created by the flagellum. They are stuck on the microvilli and collected at the base of the collar, where they are phagocytosed. The cell body can be protected by a theca which takes the name of lorica when it contains silica.The cells are surrounded by a glycocalyx, i.e. a fibrillar structure similar to a simple extracellular matrix. This glycocalyx can thicken and the species are then protected by an organic theca. A particular group uses a siliceous structure in the form of a woven basket as protection: the lorica (Figure 89). This last group is monophyletic and marine. It has silica transporters related to those of diatoms (Bacillariophyta), which also produce a siliceous protection, the frustule. During their division, the species possessing a lorica emit projections which are probably pseudopodia, showing that these organisms still have the capacity to make amoeboid movements. Some species, such as Codosiga botrytis, can form resistance cysts (Figure 89). These appear to be very resistant, because cysts preserved in the permafrost at an age of about 40,000 years may be awakened. They gave birth to individuals similar in every way to today’s individuals.
Many species of Choanoflagellata actually have the ability to differentiate several cell types based on external constraints (Figure 90). The transition from the unicellular to the colonial form has been extensively studied in Salpingoeca rosetta, which seems to become the model system for this group. It has been possible to show that the presence of sulfonolipids at concentrations as low as 10-15 M is the signal which leads to the formation of colonies. The colonies do not result from the aggregation of cells, but from their division and incomplete separation. Their formation requires a type C lectin which localizes in the center of the colony. Sulfonolipids are secreted by Algoriphagus machipongonensis, the bacterium which serves as prey for Salpingoeca rosetta. The formation of colonies has the effect of improving the flow of water around the organism and therefore of facilitating the harvest of food. Bacteria may therefore have participated in the development of multicellularity in animals by providing the first signaling molecules.
Our knowledge of the Choanoflagellata life cycle remains incomplete. Sexual reproduction has only been described in some isolates of Salpingoeca rosetta (Figure 90). It involves two flagellated gametes of different sizes. These are differentiated when the environment becomes impoverished, by a still unknown mechanism. The same culture gives both types of gametes, so it would seem that Salpingoeca rosetta is homothallic. Diploid, like haploid cells, can divide and appear to cause meiosis when returned to rich media, but this has not yet been demonstrated cytologically. Nevertheless, genetic recombination has been detected, suggesting a meiotic process. Choanoflagellata genomes remain small (40-50 Mb) and compact with around 10,000 genes present. As in the case of Capsaspora owczarzaki, some of these genes are orthologs of genes involved in the construction of multicellularity, such as the nervous system, suggesting that the ancestor common to Choanoflagellata and Metazoa already possessed some of the genes important to make an animal. Probably some of these genes are used to manage the differentiation and colony formation seen in Choanoflagellata and other unicellular Filozoa.
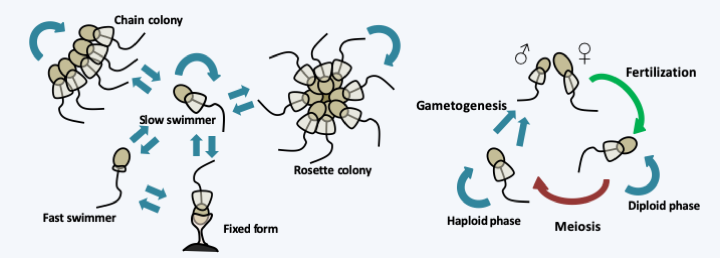
Figure 90.
Differentiation and life cycle of Salpingoeca rosetta.Metazoa
It seems surprising to talk about animals (Metazoa) in a book on protists, but it is difficult not to mention them in a chapter on Holozoa, especially since some animals, and more particularly sponges, have long been classified among protists (Figure 63) and are still considered by some biologists as intermediaries between real animals or Eumetazoa, and “protozoa”. Molecular phylogenies have so far been unable to solve the problem of the origin of animals.
[Translator’s note: Here follows a section about the evolution of Metazoa from protists, including a long section about Porifera]
Back to chapter index