Opisthokonta
Back to main indexThe Opisthokonta contains rather morphologically and biologically heterogeneous species, since it includes both Eumycota and Metazoa. However, it constitutes a phylogenetic lineage now firmly established thanks to a collection of converging phylogenies and molecular signatures. It is the eukaryotic line that produced the greatest variety of species, as the numbers of Eumycota and Metazoa species are estimated to be in the millions. In addition to these two major groups, it contains several other lineages (Figure 84), including Rozellida and amoeba Nucleariida, which are close relatives of Eumycota, and Choanoflagellata, the single-celled sibling group of Metazoa.
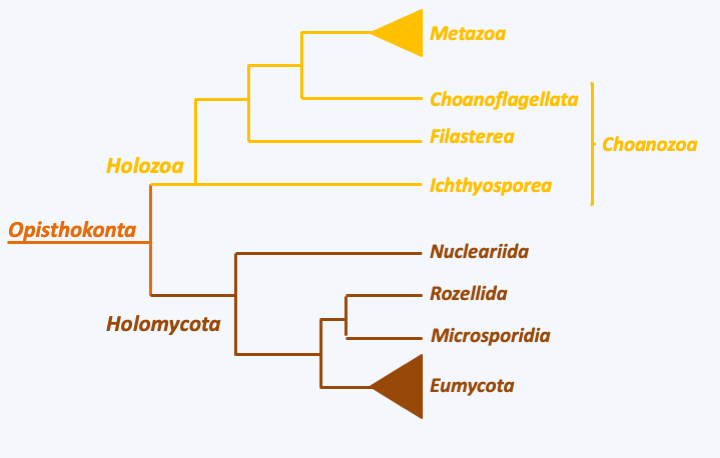
Figure 84.
Phylogenetic tree of Opisthokonta.Many characters are shared by these organisms. Thymidylate synthetase and dihydrofolate reductase are two separate enzymes whereas they are fused in all other eukaryotes except Amoebozoa. When not replaced by EFL, the translational elongation factor eEF1A, one of the most conserved proteins, has an insertion of 9 to 15 amino acids depending on the lineage, absent from other eukaryotes. Opisthokonta have the ability to synthesize chitin, which they use to strengthen the cell walls. They store their carbon reserves in the form of glycogen. In mitochondria, the UGA codon specifies tryptophan instead of stop. However, the most iconic trait that earned them their name is the presence of a single flagellum that propels their flagellated cells (Figure 85). However, at the base of the single flagellum is a pair of basal bodies, most often perpendicular to each other, betraying the biflagellar origin of the flagellar structure of Opisthokonta.
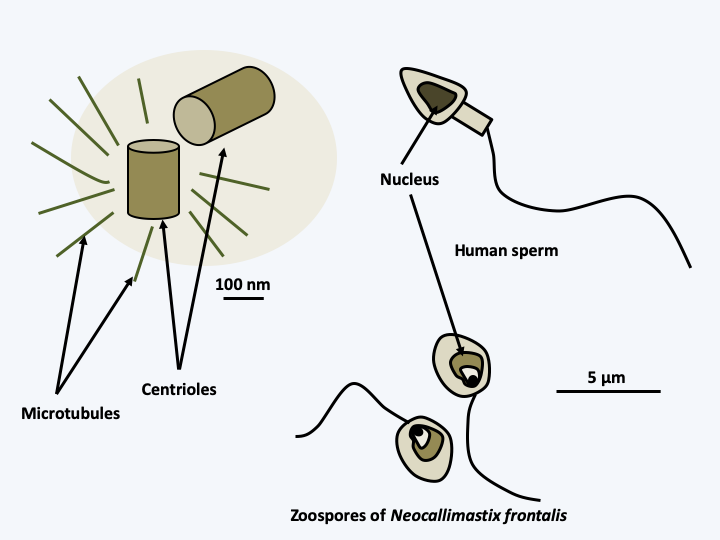
Figure 85.
Structure of the flagellum in Opisthokonts. The majority of Opisthokonts differentiate at some point in their cycle a flagellated cell with a single posterior flagellum. In animals, these are the sperm and the zoospore in 'chytridiomycete' fungi, such as Neocallimastix frontalis. In both cases, these cells are involved in dispersal/reproduction and are no longer able to feed. Despite the presence of a single axoneme, the base of the flagellum contains two basal bodies (centrioles), a trace of the biflagellar past of the Opisthokonts. In the majority of species the two centrioles are perpendicular, but there are exceptions in certain flagellate fungi.The phylogenies establish the existence of two major evolutionary lineages of Opisthokonta, that of the Holozoa leading to animals, and that of the Holomycota to “true” fungi. They show that animals are closely related to groups of unicellular protists, including Choanoflagellata, and a set of species, most of which are parasites, grouped together in Filasterea and Ichtyosporea. Eumycota fungi are related to Rozellida, which appear to feed by phagotrophy, and to Microsporidia whose parasitism has resulted in such reduction that these organisms have long been considered as intermediaries between eukaryotes and prokaryotes. The closest relatives to them are amoeba, Nuclearia and Fonticula. This assemblage would have separated from the other eukaryotes around 1000 Ma ago. The separation between the Holozoa and the Holomycota would have occurred shortly after and that between the Choanoflagellata and the Metazoa around 800 Ma ago.
In addition to the sequencing programs dedicated to Metazoa and Eumycota, an effort aimed at establishing the genomic sequences of several unicellular Opisthokonta organisms should make it possible to better understand their evolution and more particularly how multicellularity is established in Metazoa and Eumycota. The fact that these organisms probably diverged earlier from the common Opishokontan ancestor than true animals and fungi gives us some idea of the structure of the ancestor. The presence among these simple species of flagellate groups and amoeboid groups confirms that this ancestor must have been a phagotroph capable of oscillating between an amoeba form and the flagellate form.
Back to chapter index