Amoebozoa
Back to main indexJump to section:
Introduction
The second great line of Amorphea, apart from Opisthokonta, is the branch of Amoeboza. It has about 5,000 species which for the most part are phagotrophic amoebae moving with the help of lobed pseudopods, that is to say having a rather cylindrical shape and with two compartments: an internal granular or endoplasm and a peripheral hyaline or ectoplasm (Figure 184). Amoebae can be naked or protected by tests of varying shapes and compositions. The tests or thecae are protection of variable composition which surrounds the cells. There are species capable of differentiating flagella, the number of which ranges from one in Phalansterium to several dozen in Multicilia. However, the flagellated forms usually have two flagella. Recurrently, the Amoebozoa evolved forms of dispersion via spores mounted on small “feet”. In several lineages, this culminates in the differentiation of sorocarps, the sporophores of these organisms, multicellular. Like Opisthokonta, Amoebozoa have two distinct genes encoding dihydrofolate reductase and thimidilate synthetase. These organisms are very common in freshwater and soils. Most are phagotrophs feeding on bacteria or protists. Some are parasites, one of which is very dangerous for humans, Entamoeba histolytica.
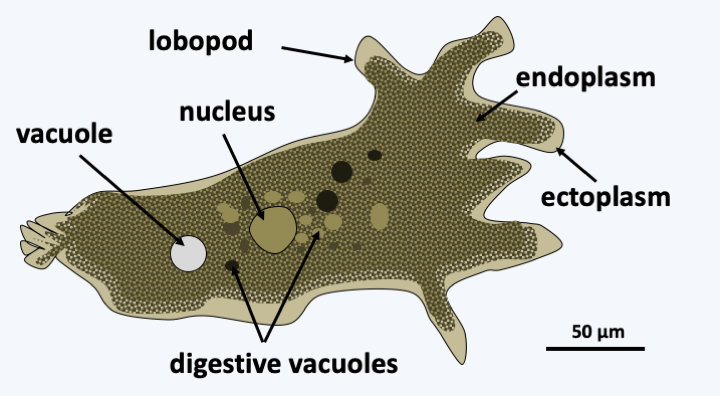
Figure 184.
Amoeba proteus, a typical Amoebozoa.Molecular phylogenies separate the Amoebozoa into two sub-branches, the Conosa and the Lobosa (Figure 185). Classification is difficult, because of a lack of morphological characters along with numerous convergent evolutions relating to the appearance of multicellularity, or the form and composition of the tests. Synapomorphies for each of the two sub-branches are rare. Conosa can still differentiate flagella while Lobosa live only as amoeba. For each of the sub-branches, the independent evolutionary lines are numerous and only the most important are indicated in Figure 185.
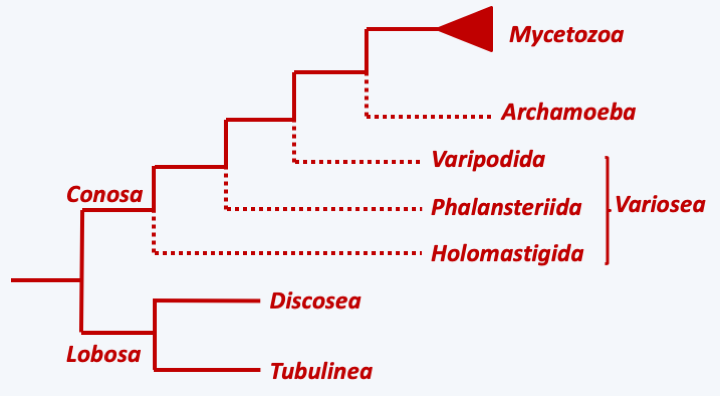
Figure 185.
Phylogeny of Amoebozoa.Lobosa
The Lobosa subphylum currently contains two classes, Discosea and Tubulinea. Amoeba in this group are phagotrophs that live mainly in muddy sediments and moist soils. They have various morphologies and tests (Figure 186). Molecular phylogenies have made it possible to better understand their evolution. The species protected by tests are grouped within Tubulinea with naked amoebae, the best known of which is Amoeba proteus (Figure 184), while Discosea mainly contain naked amoebae. Many convergences have taken place in the composition of the test which makes obsolete the old classification which was based in part on this characteristic. The new classification has made it possible to determine that the differences in the formation and morphology of the pseudopods, as well as the shape of the test and especially that of the orifice through which the pseudopods emerge, are important characters of differentiation of the different orders and families. In Lobosa, an aggregative multicellularity appeared only once: in the Copromyxa, which are of the Tubulinea class (Figure 187).
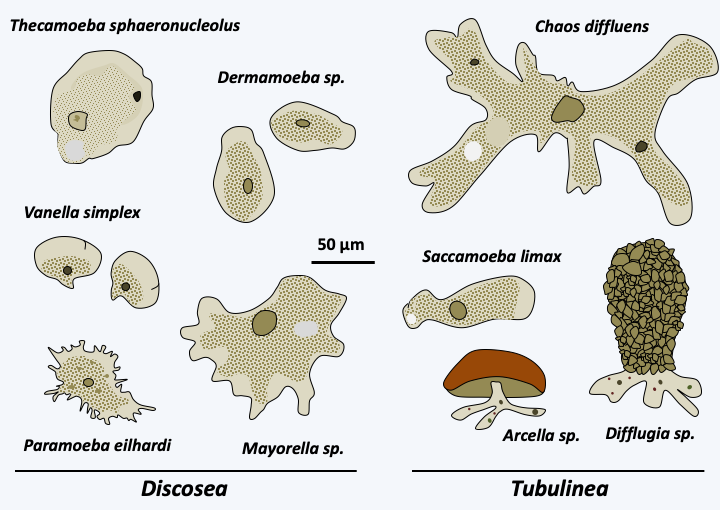
Figure 186.
Diversity of amoebae of the Lobosa sub-phylum.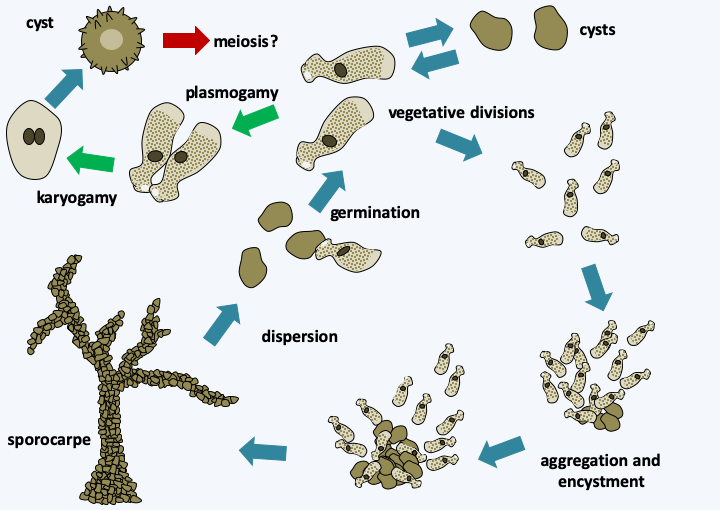
Figure 187.
Copromyxa protea life cycle. The asexual cycle of this organism passes through the formation of a multicellular sorocarp. A sexual cycle is though to be present but meiosis remains to be observed.Conosa
The subphylum of Conosa contains three classes: Variosea, Archamoeba and Mycetozoa. The first, that of the Variosea, could be paraphyletic as shown inFigure 185. The members of the group are phagotrophs that live in fresh water and moist soils. It encompasses three orders, two of which contain Amoebozoa living only or mainly in the form of flagellates (Figure 188). The most basal, that of Holomastigida, contains Multicilia, “flagellates” carrying more than twenty flagella. The order Phalansteriida contains the Phalansterium which are flagellates carrying a single flagellum. One species, Phalansterium filosum, is transiently capable of amoeboid movements. The pseudopods it produces are rather thin. Amoebae of the order Varipoda have the same fine pseudopodia.
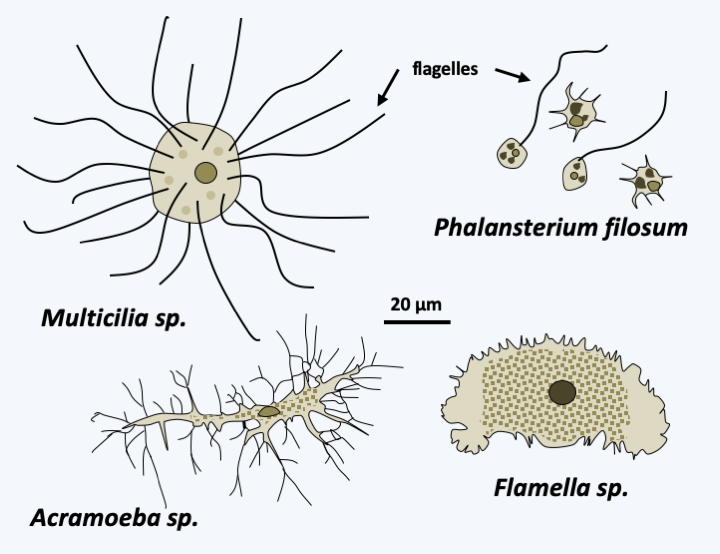
Figure 188.
Some representatives of the Variosea class.Archamoeba
The Archamoeba class contains amoeba which live anaerobically and therefore no longer breathe. This transition of habitat was accompanied by regression of the mitochondria until it persisted in some species only as a mitosome. Most Archamoeba have a flagellum (Figure 189), but this is not very active because the dyneins allowing movement are incomplete. In some species, flagellar disorganization is even more pronounced with, for example, basal bodies having only doublets instead of triplets of microtubules. In the Entamoeba, the flagellum is absent and left no trace. These phagotrophs live either in sediments loaded with organic matter, or in commensal or parasitic association with animals. This is the case with Entamoeba gingivalis (Figure 189) which lives in our mouth, as well as Entamoeba intestinalis, and these species disappear in our intestines as commensals. Entamoeba histolytica, on the contrary, is a pathogen which can cause serious fatal dysentery. Usually the infection is mild because it remains localized in the intestine. However, Entamoeba histolytica can enter the blood vessels and cause bloody diarrhea. Once the intestinal barrier is crossed, the amoeba can colonize other parts of the body such as the lungs, brain and especially the liver where it will cause abscesses. The infection then becomes very dangerous and even fatal. It is estimated that 10% of the human population is infected with Entamoeba histolytica which is the third leading cause of mortality among parasites, causing approximately 50,000 to 100,000 deaths per year. They have a life cycle devoid of sexuality (Figure 190).
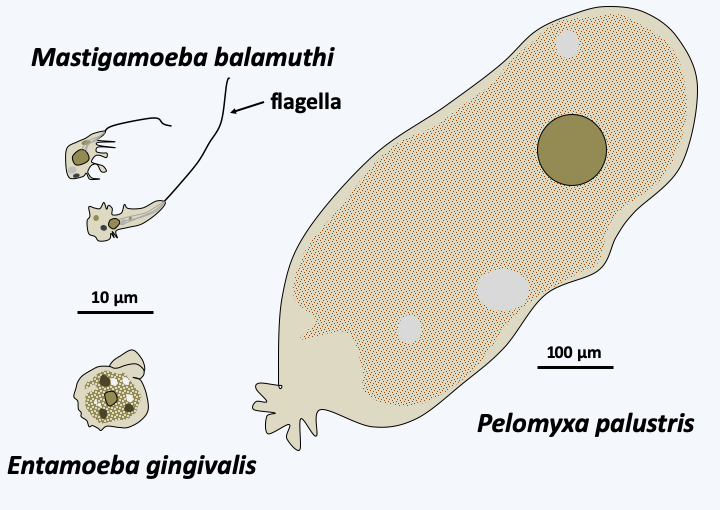
Figure 189.
Some representatives of the Archamoeba class.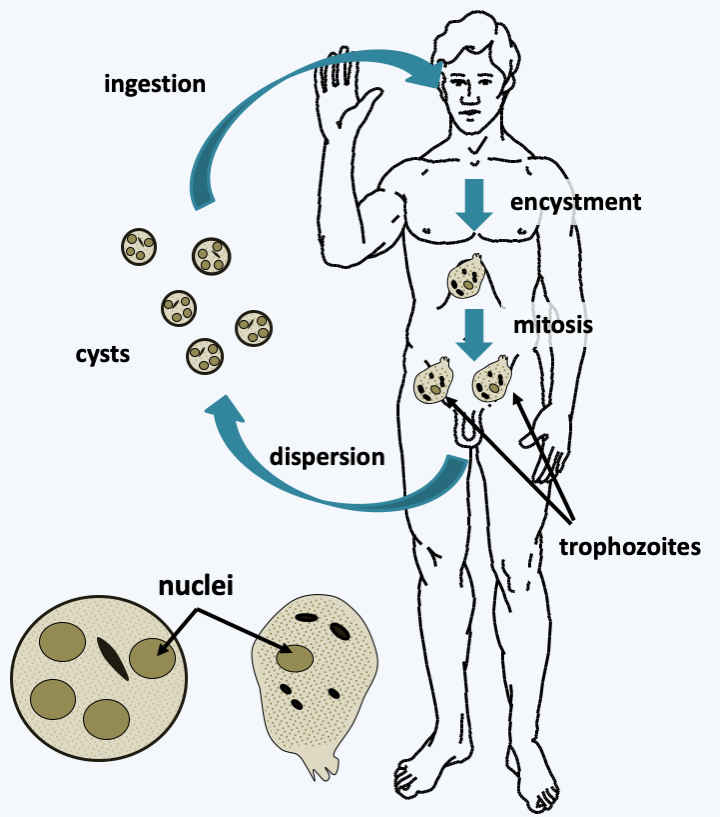
Figure 190.
Entamoeba histolytica cycle. Quadrinucleate cysts are the infectious forms, although they provide only relative protection and are quite fragile. After entering the intestine, the digestive juices allow the emergence of trophozoites. These divide by binary fission and feed on intestinal debris. Once in the blood, Entamoeba histolytica is able to engulf erythrocytes. New cysts are released when the stool is passed.Mycetozoa
The last line of Amoebozoa, the Mycetozoa superclass, has undergone several name changes because the phylogenetic position of the organisms it contains has long been a mystery. Indeed, a good part of its members has a phagotrophic phase, a rather “animal-like” characteristic, but disperses by building multicellular sporophores called sorocarps like fungi! Initially, called Mycetozoa (= animal fungi), they were then known for over a hundred years under the name Myxomycota (gelatinous fungi; slime molds), only to recently become Mycetoza again. All of them have a phagotrophic trophic phase, which disqualifies them from being fungi! Molecular analyzes have complicated their classification by showing that many convergences have taken place during the evolution of the group, and that the old classification based on the type of development of the sporophore is obsolete. Some species formerly classified as Mycetozoa are not even Amoebozoa! Some joined the Nucleariida, like Fonticula alba, part the Discoba, like the Acrasis and Pocheina, and part the Rhizaria like Guttulinopsis, others, like the Copromyxa, joined other classes of the Amoebozoa. Traditionally, mycologists/protozoologists distinguished three types of development for group members (Figure 191). The “protostelid” type lives mainly in the form of amoeba, possibly in the form of small plasmodia having a few nuclei, and to disperse from sorocarps carrying one or a few spores. Unfortunately, this type of development has been “invented” many times in Amoebozoa, resulting in the old class of Protostelida being polyphyletic with representatives in virtually all classes of Amoebozoa! The “dictyostelid” (Read more here)type lives in the form of amoeba which will group together and give rise to a multicellular structure by aggregative multicellularity, the sorogen. This structure may be devoid of movement, but in some species it can move independently. It is then called pseudoplasmodia. As it resembles a slug, the name limax is also sometimes used. Note that aggregative multicellular organisms that are not Mycetozoa, such as Fonticula alba, Acrasis/Pocheina or Copromyxa do not differentiate pseudoplasmoda. This type of development seems to have evolved only once in the Dictyosteliida. The third type called “myxogastrid” is a complexification of the life cycle of certain protostelids. Species that have adopted this type first live as free amoebae, which later form a giant plasmodium. This often follows fertilization between sexually competent amoebae, but since there are homothallic strains, it can occur in the absence of plasmogamy between different amoebae. The plasmodium eventually gives rise to multicellular fruiting bodies, sometimes of large size, superficially resembling small fungi. This type of development has also been evolved only once, in Myxogastrea. Recently another type of development, that of “hyperamoeba” (Figure 191) has also been shown to be present in these Myxogastrea. This development is simpler, with three forms: amoeba, flagellates and cysts. It is currently not yet clear whether this is a regression with loss of the ability to form plasmodia and sorocarps, or if part of the life cycle of these Hyperamoeba has not yet been observed. All of the evolutionary convergences make the classification of Mycetozoa incomplete and complex (Figure 192). Molecular phylogenies differ greatly depending on the genes chosen to construct them and it could for example be that the other two classes of Variosea and Archamoeba are in fact mixed with that of Mycetozoa. Only the Dictyosteliida and Myxogastrea classes will be developed here, because data on the forty or so “protostelid” species are scarce and incomplete.
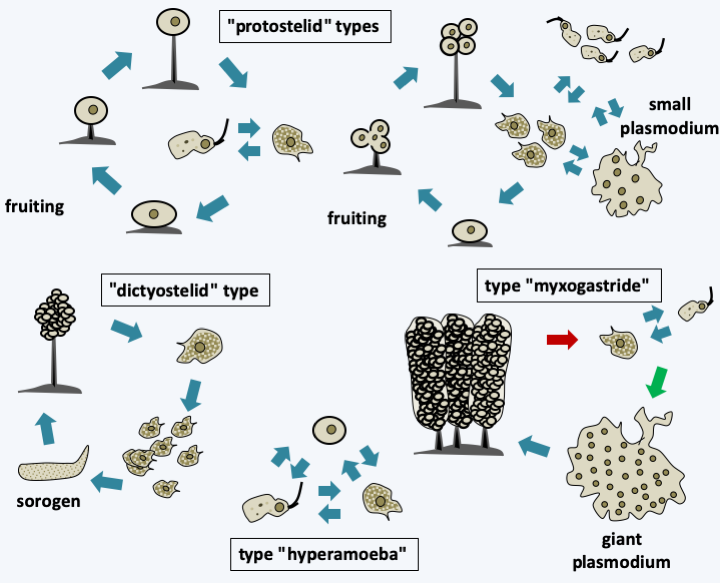
Figure 191.
Types of development formerly recognized in Mycetozoa.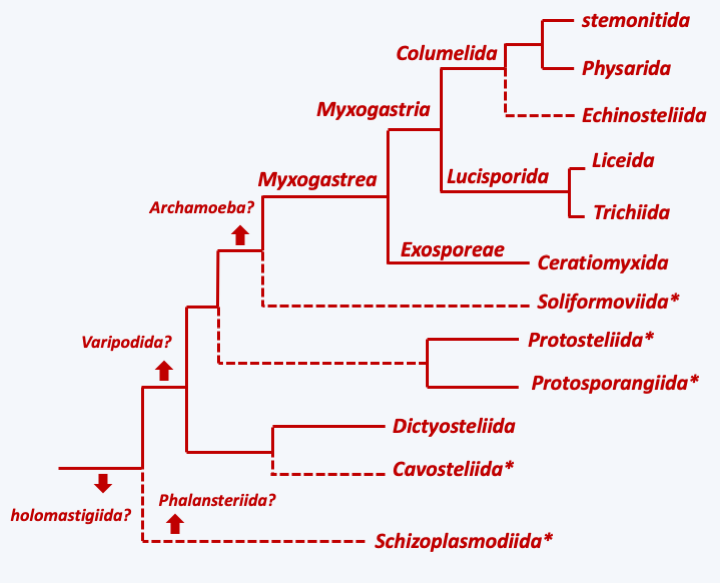
Figure 192.
Phylogeny of Mycetozoa. The arrows indicate the possible branching points of the different lines of Variosea and Archamoeba. * indicates a 'protostelid' type development.Dictyosteliida
More than a hundred species of Dictyosteliida are currently described. They are commonly found all around the world in the decaying leaf mats of deciduous forests, where they feed on bacteria by phagotrophy. Under conditions of ad libitum nutrition, amoebae can divide every 3 hours. In the laboratory, it is possible to cultivate them on a complex but defined axenic medium. The generation time is then 9 hours. The model species of the group is Dictyostelium discoideum (Figure 52 and Box 19). Different species of Dictyosteliida differ in the shape of sorocarps (Figure 193), their ability to form cysts and limax, or the molecules that serve to trigger the aggregation movement of amoebae. While cAMP plays this role in Dictyostelium discoideum and related species, in other species it is held by folic acid, a modified dipeptide called glorin, or neopterin, a degradation product of GTP. Sequence analyzes show that Dictyosteliida species began to diverge about a billion years ago. Genetically, they are therefore more diverse than animals, which is reflected in the very varied modalities of their development!
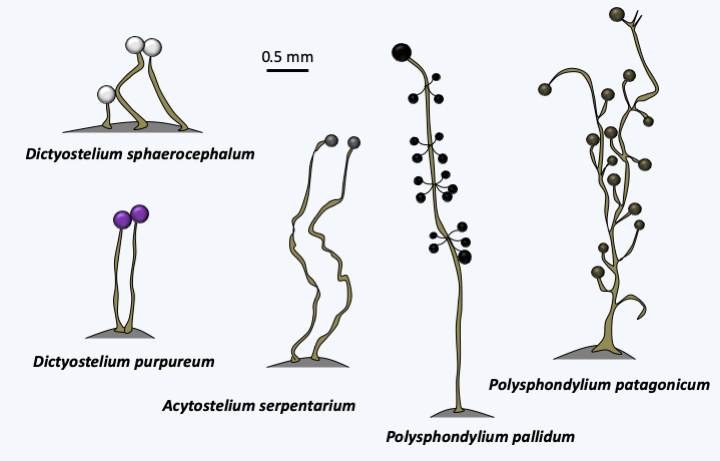
Figure 193.
Sorocarps of various Dictyosteliida.Myxogastrea
With nearly a thousand species described, and numerous new discoveries each year, the Myxogastrea class is a line of Amoebozoa that has experienced great evolutionary success. Their typical development is of the “myxogastrid” type (Figure 191) with a unicellular haploid phase and a diploid plasmodial phase. The plasmodia differentiated by these organisms are characteristic and have extraordinary properties which are of great interest to scientists (Box 20). Two subclasses are differentiated based on how their spores are produced. This distinction is supported by molecular phylogenies. The three known species belonging to the subclass Exosporeae produce their spores on the surface of the fruiting body (Figure 194). The spores are therefore directly in contact with the external environment. Of the three species, all of which belong to the order Ceratiomyxida and the genus Ceratiomyxa, one, Certiomyxa fruticulosa (Figure 194) lives in temperate forests while the other two are found in tropical forests. They all differentiate from aphanoplasmods (Box 20). Molecular analyzes suggest that Ceratiomyxa fruticulosa is a morphospecies which hides a great genetic diversity and therefore probably several species in the genetic sense.

Figure 194.
Ceratiomyxa fruticulosa. This Exosporeae is very common in temperate forests. It produces its spores externally.Unlike Exosporeae, species in the Myxogastria subclass produce their spores in sorocarps covered with a protection, the peridium. This sorocarp can take on varying shapes and sizes ranging from a few hundred microns to more than ten centimeters in diameter (Figure 195). The most common form has a disc, or hypothallus, attaching to the substrate, a stem or stipe, and a spore mass or sporocyst at the top (Figure 196). The whole thing therefore looks like a small mushroom with a stem and a globular cap. If the sorocarp looks like a fungus, it is because it performs the same function of allowing efficient dispersal of the spores in the air environment. In some species, the spores enclosed in the peridium can be supported by a structure which prolongs the stipe, the collumella. There is also sometimes a fibrillar structure, the capillitium, inside the sporocyst, which allows ejection of the spores after the peridium dries out. The model species of the group is Physarum polycephalum, in which laboratory techniques have been well mastered (Figure 197). This species is tetrapolar heterothallic with a matA locus possessing several alleles; only crosses between strains differing at the matA locus lead to the formation of the plasmodium. The other locus, matB, has a qualitative effect. It is also multiallelic and crosses between compatible strains are more than 1,000 times effective than between incompatible strains. This locus controls cell fusion, while matA regulates plasmodium formation. A third triallelic locus, matC, influences the efficiency of crosses at different pHs. Many variations exist around this life cycle. Some species always remain diploid and others always haploid. Other species can use several types of cycles, homothallic or heterothallic, and we do not yet understand their determinism.

Figure 195.
Some sorocarps of Myxogastria. From left to right and top to bottom, Enerthenema intermedium, Stemonitis fusca, Arcyria incarnata, Leocarpus fragilis, Lycogala epidendron and Fuligo septica. The first species differentiates from sorocarps that are barely visible to the naked eye, while in the last they are several centimeters in length.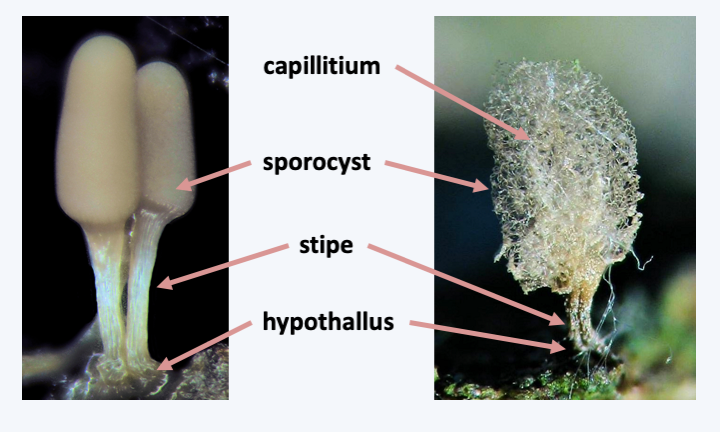
Figure 196.
Structure of a typical sorocarp of Myxogastria.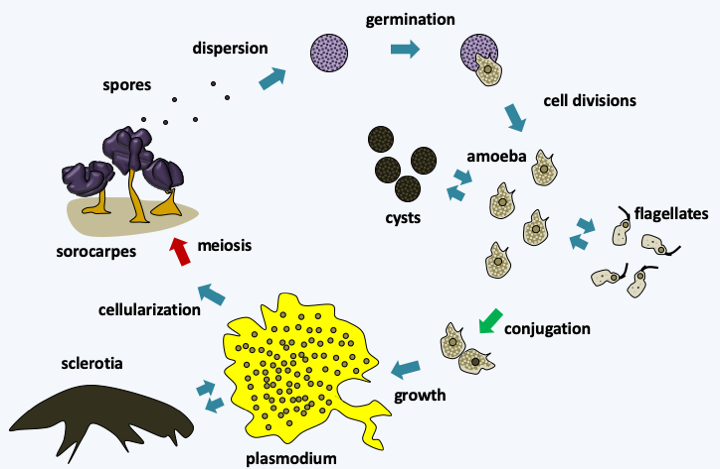
Figure 197.
Physarum polycephalum life cycle. The cycle begins with the dispersion of spores which germinate into to one or more rarely two cells when the humidity conditions are right. If the environment is rather dry, amoeba are released, if the environment is humid, biflagellate cells appear. These cells change from one form to another depending on the availability of water or can differentiate into cysts if conditions are adverse. The meeting of two sexually compatible cells gives rise to a plasmodium. This can reversibly differentiate into a sclerotia or macrocyst, if conditions get too dry. When nutrients are depleted in the medium, the plasmodium migrates to the surface of its substrate and cellularizes. Concomitantly, the cells undergo meiosis and the whole differentiates into sorocarps, where spores form. A plasmodium can, depending on its size, give rise to a few sorocarp or several hundred. The very resistant spores allow survival in the environment.One trait that appears to be shared by Myxogastria is the presence of RNA editing in the mitochondria. Most editions are insertions of C, occasionally insertions of T, AA, AU, CU, GU and GC. There may also be substitutions of C by U. The editing sites are approximately uniformly spaced 25 nucleotides apart and are localized in mRNAs, some tRNAs and rRNAs.
The traditional classification, based mainly on the morphology of the sorocarp and the ornamentation and color of the spores but also on the type of plasmodia (phanero- or aphanoplasmoda), recognized 5 orders of Myxogastrea (Figure 192). Sorocarps may be sessile or stem mounted (Figure 195, Enerthenema intermedium, Stemonitis fusca, and Leocarpus fragilis), and may or may not have columella or capillitium. They can be grouped together in the form of a “pseudo-aethalium”, the aethalium-like sorocarps in turn resembling a flattened cushion (Figure 195, Lycogala epidendon and Fuligo septica which presents an aethalium in formation). Molecular data show that there are many convergences and that this classification needs to be reviewed.
Like the Dictyosteliida, the Myxogastrea are rather discreet and have little connection with humans. There are no parasites or species of pharmaceutical interest. They are nevertheless extremely common in forests and humid undergrowth all around the earth, but they can also inhabit restricted biotopes such as the species that live on the edges of melting glaciers or those that live in deserts. Like the Eumycota, they are easy to distinguish at the time of their fruiting. To observe them, all you need to do is collect tree bark or pieces of dead wood, which must then be left to soak overnight in water. The samples are then kept in constant humidity. After incubation for several days, plasmodia and/or sorocarps are easily observed with a magnifying glass or the naked eye. Sorocarps of Dictyosteliida can also be obtained in this way; the best source being forest soil samples. “Raising” Myxogastrea plasmodia is very easy because you just need to feed them with oatmeal!
Back to chapter index